

{kind=link}
{kind=link}
{kind=link}
{kind=link}
尿路上皮癌相关1基因通过竞争性抑制 miR-18a增强乳腺癌细胞的他莫昔芬治疗耐药
[李秀楠△  , 刘爱蕙, 唐欣, 任宇]
, 刘爱蕙, 唐欣, 任宇]
, 刘爱蕙, 唐欣, 任宇]
|
|


目的 探讨尿路上皮癌相关1(urothelial carcinoma-associated 1, UCA1)基因及 miR-18a在雌激素受体(estrogen receptor, ER)阳性的乳腺癌细胞他莫昔芬耐药中的作用和调控机制。方法 通过反转录实时定量PCR测定乳腺癌细胞中 UCA1和 miR-18a的表达,用双荧光素酶报告实验验证 miR-18a和 UCA1 3'UTR区域结合。在他莫昔芬敏感的MCF-7细胞中过表达 UCA1或敲减 miR-18a,在他莫昔芬耐药的LCC9和BT474细胞中敲减 UCA1或过表达 miR-18a,CCK-8方法测定细胞活力,软琼脂克隆形成实验测定细胞克隆形成能力,流式细胞术测定细胞周期。结果 他莫昔芬耐药的LCC2、LCC9和BT474细胞中, UCA1表达量显著高于他莫昔芬敏感的MCF-7细胞。MCF-7细胞在0.1 μmol/L 他莫昔芬处理后, UCA1的表达水平显著升高。 UCA1过表达提高了MCF-7细胞在他莫昔芬处理后的活力,而 UCA1 siRNA则显著降低了LCC9和BT474细胞在他莫昔芬处理后的活力。在MCF-7细胞中,相对于质粒对照+他莫昔芬组, UCA1+他莫昔芬组的克隆数目多19.0%,克隆平均大小增加29.0%,G1期细胞比例减少7.3%,S期细胞增加6.7%。在BT474细胞中,相对于siRNA对照+他莫昔芬组, si-UCA1+他莫昔芬组的克隆数目少54.0%,克隆平均大小减少42.0%,G1期细胞比例增加9.0%,S期细胞减少6.2%。 UCA1有一个 miR-18a结合位点,敲减 miR-18a提高了MCF-7细胞在他莫昔芬处理后的活力,过表达 miR-18a则显著降低了BT474细胞在他莫昔芬处理后的活力。在MCF-7细胞中,相对于对照+他莫昔芬组, miR-18a抑制+他莫昔芬组的克隆数目多15.0%,克隆平均大小增加33.0%,G1期细胞比例减少8.8%,S期细胞增加5.3%。在BT474细胞中,相对于对照+他莫昔芬组, miR-18a过表达+他莫昔芬组的克隆数目少47.0%,克隆平均大小减少25.0%,G1期细胞比例增加13.3%,S期细胞减少7.9%。结论 UCA1可以通过竞争性抑制 miR-18a增强乳腺癌细胞的他莫昔芬治疗耐药。
Objective: To investigate how urothelial carcinoma-associated 1 ( UCA1) and miR-18a modulates acquired tamoxifen resistance and the relevant mechanisms in estrogen receptor (ER) positive cancer cells.Methods: qRT-PCR was performed to detect UCA1 and miR-18a expression in breast cancer cells. Dual luciferase assay was performed to detect the binding between miR-18a and UCA1 3'UTR. Tamoxifen sensitive MCF-7 cells were transfected with UCA1 expression vector or miR-18a inhi-bitors. Tamoxifen resistant LCC9 and BT474 cells were transfected with UCA1 siRNA or miR-18a mi-mics. CCK-8 assay was performed to detect cell viability. Soft agar assay was performed to assess cell colony formation. Flow cytometric analysis was performed to check cell cycle distribution.Results: UCA1 was significantly upregulated in tamoxifen resistant LCC2, LCC9, and BT474 cells than in tamoxifen sensitive MCF-7 cells. UCA1 expression was significantly upregulated in MCF-7 cells after treatment with 0.1 μmol/L tamoxifen. UCA1 overexpression enhanced cell viability of MCF-7 cells after tamoxifen treatment, while UCA1 siRNA significantly suppressed viability of LCC9 and BT474 cells after tamoxifen treatment. In MCF-7 cells, compared with vector control+tamoxifen group, the average cell colony number and colony size of the UCA1+tamoxifen group was 19.0% more and 29.0% larger respectively, while the proportions of the cells in G1 phase and in S phase were 7.3% lower and 6.7% higher respectively. In BT474 cells, compared with siRNA control+tamoxifen group, the average cell colony number and colony size of the si-UCA1+tamoxifen group were 54.0% less and 42.0% smaller respectively, while the proportions of the cells in G1 phase and in S phase were 9.0% higher and 6.2% lower respectively. UCA1 directly interacted with miR-18a and reduced its expression in ER positive breast cancer cells. Knockdown of miR-18a increased viability of MCF-7 cells after tamoxifen treatment, while miR-18a overexpression significantly reduced viability of BT474 cells after tamoxifen treatment. In MCF-7 cells, compared with miRNA inhibitor control+tamoxifen group, the average cell colony number and colony size of the miR-18a inhibitor+tamoxifen group were 15.0% more and 33.0% larger respectively, while the proportions of the cells in G1 phase and in S phase were 8.8% lower and 5.3% higher respectively. In BT474 cells, compared with miRNA control+tamoxifen group, the average cell colony number and colony size of the miR-18a mimics+tamoxifen group were 47.0% less and 25.0% smaller respectively, while the proportions of the cells in G1 phase and in S phase were 13.3% higher and 7.9% lower respectively.Conclusion: UCA1 can increase tamoxifen resistance of ER positive breast cancer cells via competitively inhibiting of miR-18a.
乳腺癌是女性最常见的恶性肿瘤之一, 也是女性肿瘤患者中死亡率第二位的肿瘤[1]。在所有乳腺癌肿瘤中, 大约70%为雌激素受体(estrogen receptor, ER)阳性[2, 3], 因此, 他莫昔芬作为雌二醇竞争性拮抗剂, 被广泛用于ER阳性的早期乳腺癌患者术后的辅助治疗[4]。虽然他莫昔芬的临床使用显著提高了患者的生存期, 但是乳腺癌细胞通常会在一段时间治疗后出现他莫昔芬耐药, 从而导致肿瘤复发和他莫昔芬治疗失败[4]。他莫昔芬的耐药机制十分复杂, 很多分子机制目前仍然未知。
长链非编码RNA(long noncoding RNA, lnc-RNA)是一类转录本长度超过200个核苷酸的RNA分子, 并不具有编码蛋白的能力[5]。最近的研究发现, lncRNA可以参与许多信号通路的调控, 从而促进或者抑制肿瘤[6, 7, 8], 也有研究报道, lncRNA在乳腺癌的内分泌治疗中可能起到比较关键的调控作用[9, 10], 比如在他莫昔芬敏感的ZR-75-1细胞中过表达乳腺癌抗雌激素药物耐药性基因4(breast cancer anti-estrogen resistance 4, BCAR4)可以部分解除他莫昔芬对细胞增殖的抑制作用[10], 因此, BCAR4也被认为是乳腺癌他莫昔芬耐药的标志物之一[9, 11]。另外一项研究发现, 长链基因间非编码RNA1016和00160(LINC01016和LINC00160)是直接的ERα 转录调控靶点。LINC01016和LINC00160的表达与ERα 的表达呈正相关, 且LINC01016和LINC00160的高表达预示了患者的不良预后[12]。在乳腺细胞中敲减LINC00160可以显著降低细胞增殖水平[12]。lncRNA HOTAIR上调可以促进非配体依赖的ER活性并且促进他莫昔芬耐药[13]。
尿路上皮癌相关1(urothelial carcinoma-associa-ted 1, UCA1)基因是在膀胱移行上皮细胞癌中发现并命名的lncRNA[14]。UCA1有3个外显子, 编码两个转录本, 包括一个1.4 kb和一个2.2 kb的转录本, 1.4 kb的转录本包括在2.2 kb的转录本中[15]。最近的研究证实, 1.4 kb的UCA1转录本在乳腺癌中是一个原癌基因, UCA1在ER阳性及ER阴性的乳腺癌细胞中的表达均显著高于正常乳腺细胞[15, 16, 17]。功能上, UCA1可以通过与核不均一核糖核蛋白Ⅰ (heterogeneous nuclear ribonucleo protein, hnRNP Ⅰ )形成复合物来降低p27的表达[15]。UCA1也可通过分子海绵效应降低miR-143的表达来调控ER阳性乳腺癌细胞的生长和凋亡[18]。除此以外, UCA1也可以调控ER阳性MCF-7细胞的阿霉素敏感性[19], UCA1上调可以促进ER阴性的MDA-MB-231细胞上皮间质转化[16]。但是, 目前有关UCA1表达与ER阳性乳腺癌细胞的他莫昔芬敏感性关系的报道较少, 本研究旨在明确UCA1对ER阳性乳腺癌细胞的他莫昔芬敏感性调控, 进而探究下游的分子机制, 为乳腺癌精准治疗提供实验依据。
ER阳性且他莫昔芬敏感的乳腺癌细胞MCF-7, ER阳性、人类表皮生长因子受体2(human epidermal growth factor receptor-2, HER2)阳性且他莫昔芬耐药的乳腺癌细胞BT-474均购自美国ATCC细胞库。基于MCF-7诱导的他莫昔芬耐药的LCC2和LCC9细胞均购自中国科学院上海生命科学研究院细胞资源中心。肿瘤细胞用RMPI-1640培养基培养, 培养基中添加10%(体积分数)胎牛血清(fetal bovine serum, FBS)、2 mmol/L谷氨酰胺、100 U/mL青霉素和100 mg/L链霉素。
以MCF-7的总RNA为模板, 使用反转录试剂盒(Promega)进行反转录PCR获得1.4 kb的UCA1序列。UCA1的引物序列为:正向引物5'-CGCGGATCCTTTATCAGGCATATTAGCTTTAA-3', 反向引物5'-GCGAATTCTGACATTCTTCTGGACAATG-3'。把序列克隆到pcDNA3.1(+) (Invitrogen) 的EcoR Ⅰ 和BamR Ⅰ 位点之间, 重组质粒命名为pc-DNA3.1-UCA1。将重组质粒转染到MCF-7细胞中实现UCA1过表达, 转染试剂为Lipofectamie 2000 (Invitrogen)。
miR-18a模拟物 (mimics)、miR-18a抑制物 (inhibitors, IH)、UCA1小干扰RNA(small interfe-ring RNA, siRNA)和对应的阴性对照全部购自广州市锐博生物科技有限公司。在准备miR-mimics和siRNA转染实验的12 h前, 将所需细胞按30%密度接种于培养皿中。参考转染试剂Lipofectamie 2000 (Invitrogen) 说明书将miR-mimics或siRNA和转染试剂分别加入无血清DMEM培养基中, 静置5 min后混合, 再次静置20 min, 将混合好的DMEM培养基加入换为新培养基含血清的细胞培养皿中, 转染24 h后更换新培养基, 之后按需要浓度加入他莫昔芬, 处理指定时间后收集细胞。
联合使用PITA (http://genie.weizmann.ac.il/pubs/mir07/mir07_prediction.html)和RNAhybrid (http://bibiserv2.cebitec.uni-bielefeld.de/rnahybrid)来预测UCA1可能的miRNA结合位点。
按照TRIzolRNA提取试剂盒说明书提取细胞总RNA, 以细胞总RNA为模板, 应用PrimeScript® 逆转录试剂盒(TaKaRa)或TaqMan miRNA逆转录试剂盒(Applied Biosystems)将RNA逆转录为cDNA。根据qRT-PCR试剂盒说明书和基因对应的引物对cDNA进行扩增。UCA1的引物为:正向引物5'-CTCTCCATTGGGTTCACCATTC-3', 反向引物5'-GCGGCAGGTCTTAAGAGATGAG-3'。用TaqMan miRNA分析试剂盒检测miR-18a的表达, 使用U6作为内参基因。所有qRT-PCR实验均使用Applied Biosystems 7500 实时定量PCR仪进行, RNA的相对表达量使用2-Δ Δ CT进行计算。
取转染后24 h的MCF-7细胞, 调整细胞密度为3× 104个/mL, 取96孔板每孔接种100 μ L细胞悬液, 放入培养箱继续培养24 h, 之后将培养基换为含不同浓度他莫昔芬(0.1、0.5、1.0、5.0、10.0、15.0、20.0、50.0 μ mol/L)的培养基。细胞继续培养5 d后进行CCK-8检测, 检测时每孔加10 μ L CCK-8检测试剂, 放于37 ℃、5%(体积分数)CO2培养箱中孵育2 h。用酶标仪测定450 nm波长的光密度值(
将1.2%(质量分数)低熔点琼脂糖与2× 细胞培养基以体积比1 :1混合制备0.6%(质量分数)的底层琼脂, 6孔板中每个孔加0.8 mL, 4℃凝固。取对数期细胞, 胰酶消化后吹散成单个细胞悬液, 计数。将0.6%低熔点琼脂糖与2× 细胞培养基以体积比1 :1混合, 制备0.3%(质量分数)的上层琼脂, 每孔加1 mL上层琼脂和单细胞悬液(约5× 104细胞/孔), 混匀, 4℃ 5 min凝固。置于37 ℃、5% CO2培养箱中培养2周。细胞克隆用0.5%(体积分数)结晶紫染色, 计数含50个细胞以上的克隆, 计算细胞集落形成率=(克隆形成数/初始细胞数)× 100%。实验组及阴性对照组分别设3个复孔, 实验重复3次。
消化好的单细胞悬液先采用70%(体积分数)乙醇4℃固定过夜, 后用1× PBS洗涤3次。固定好的细胞用100 mg/L RNase A 37 ℃孵育30 min, 之后用10 mg/L propidium iodide (PI)混匀后4℃避光染色30 min。用流式细胞仪(FACSCaliber, USA) 488 nm激发波长检测DNA染色。实验组及阴性对照组分别设3个重复, 实验重复3次。
将含预测的野生型miR-18a结合位点的UCA1序列片段和含突变型miR-18a结合位点的UCA1片段分别插入荧光素酶报告质粒pmirGLO (Promega)的XhoⅠ 和XbaⅠ 位点之间, 重组质粒分别命名为pmirGLO-UCA1-WT和pmirGLO-UCA1-MT, 以表达海肾荧光素酶的载体pRL-TK作为内参对照, 将miR-18a mimics、mimics对照分别与荧光素酶报告载体共转染入MCF-7细胞中, 转染24 h后采用Promega公司的Dual-Lucifersae Reporter Assay System进行样品Lucifersae活性检测。
所有数据采用SPSS 18.0软件进行分析, 以均数± 标准差表示, 对多组数据采用单因素方差分析和Student-Newman-Keuls (SNK)法两两比较分析, 两组间数据比较采用t检验, P< 0.05为差异有统计学意义。
实时定量PCR结果显示, 他莫昔芬耐药的LCC2、LCC9和BT474细胞中UCA1表达量显著高于他莫昔芬敏感的MCF-7细胞(图1A)。MCF-7细胞在0.1 μ mol/L他莫昔芬处理后, UCA1的表达水平也显著升高, 第72小时升高了(7.7± 0.9)倍, 第96小时升高了(8.6± 1.1)倍(图1B)。
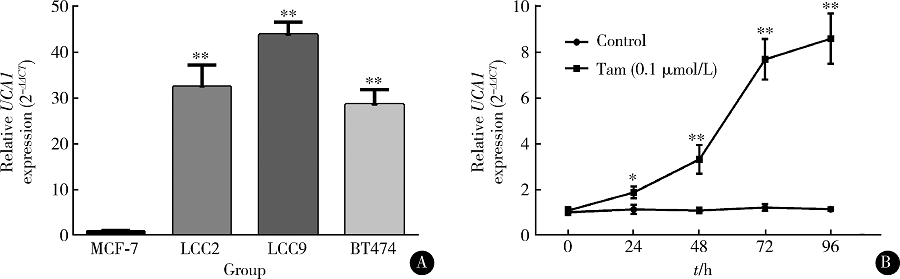 | 图1 ER阳性乳腺癌细胞中的lncRNA UCA1在他莫昔芬处理后表达显著升高Figure 1 lncRNA UCA1 is significantly upregulated after tamoxifen treatment in ER-positive breast cancer cellsA, qRT-PCR analysis of UCA1 expression in MCF-7, LCC2, LCC9, and BT474 cells; B, qRT-PCR analysis of UCA1 expression in MCF-7 after tamoxifen (0.1 μ mol/L) treatment at indicating time points up to 96 h. * P< 0.05, * * P< 0.01, compared with MCF-7 group or control group. Tam, tamoxifen. |
为了研究UCA1的他莫昔芬敏感性调节作用, 将MCF-7细胞转染UCA1过表达载体pcDNA3.1-UCA1, LCC2和BT474细胞转染UCA1 siRNA。细胞活力实验结果显示, UCA1过表达显著提高了MCF-7细胞在不同浓度他莫昔芬处理后的细胞活力(图2A), 而UCA1 siRNA则显著降低了LCC9和BT474细胞在他莫昔芬处理后的活力(图2B、C)。细胞克隆形成实验结果显示, 在MCF-7细胞中过表达UCA1, 可以部分解除他莫昔芬抑制克隆形成作用。相对于质粒对照+他莫昔芬组, UCA1+他莫昔芬组的克隆数目多19.0%, 克隆平均大小增加29.0%(图2D、E)。相反, 在BT474细胞中抑制UCA1可以显著增加他莫昔芬抑制克隆形成作用, 相对于siRNA对照+他莫昔芬组, si-UCA1+他莫昔芬组的克隆数目少54.0%, 克隆平均大小减少42.0% (图2D、E)。通过流式细胞术分析细胞周期结果表明, 在MCF-7细胞中过表达UCA1可以部分解除他莫昔芬诱导的G0/G1细胞周期阻滞。相对于质粒对照+他莫昔芬组, UCA1+他莫昔芬组G1期细胞比例减少7.3%, S期细胞增加6.7% (图2F、G)。相反, 在BT474细胞中抑制UCA1可以显著增加他莫昔芬诱导的G0/G1细胞周期阻滞, 相对于 siRNA对照+他莫昔芬组, si-UCA1+他莫昔芬组G1期细胞比例增加9.0%, S期细胞减少6.2% (图2F、G、H)。
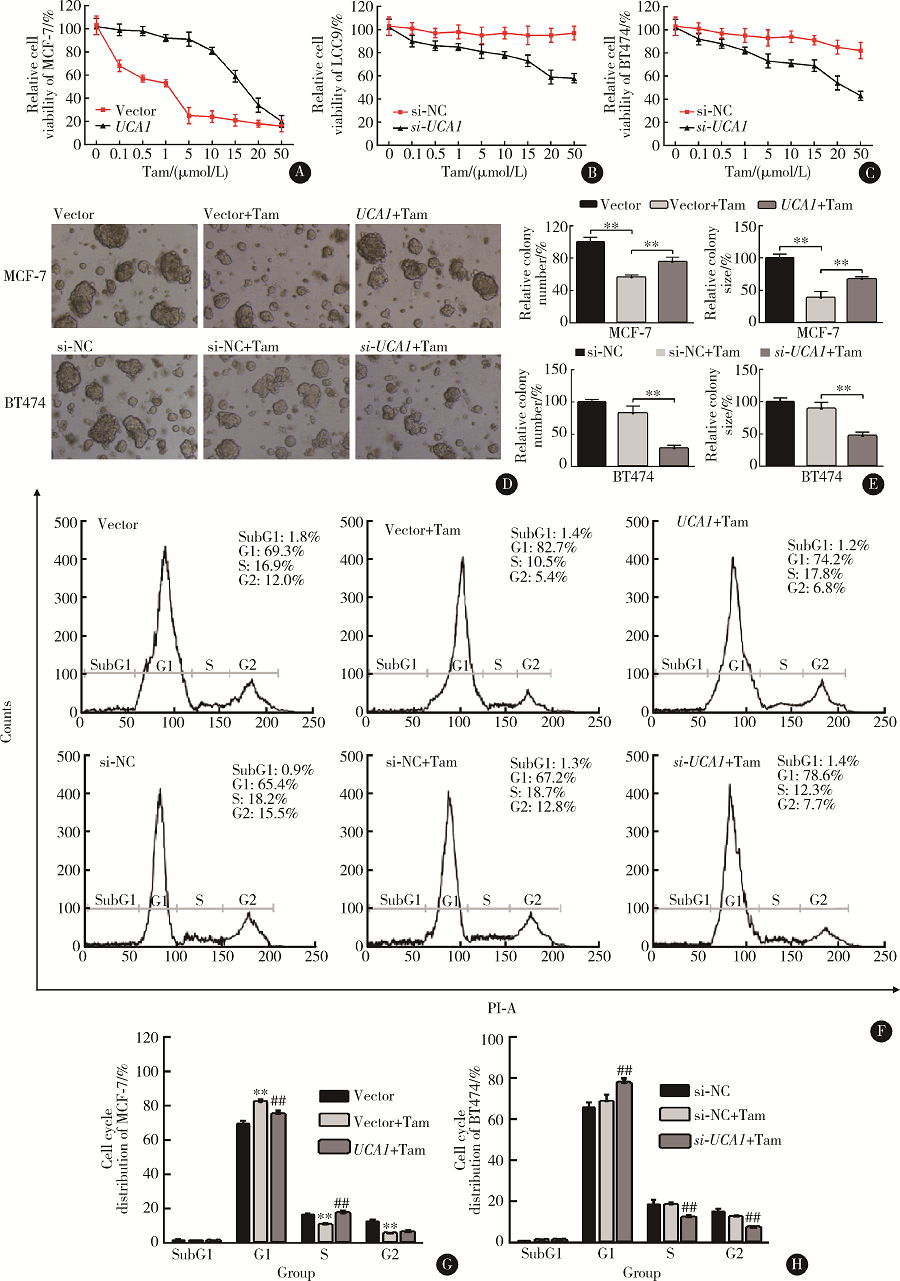 | 图2 lncRNA UCA1可以降低ER阳性乳腺癌细胞的他莫昔芬敏感性Figure 2 lncRNA UCA1 upregulation results in acquired tamoxifen resistance in ER-positive breast cancer cellsA-C, CCK-8 assay of cell viability of MCF-7 cells with or without UCA1 overexpression (A), LCC9 (B) and BT474 cells (C) with or without UCA1 knockdown after treatment with varying concentrations of tamoxifen (0.1, 0.5, 1.0, 5.0, 10.0, 15.0, 20.0, 50.0 μ mol/L) for 5 days; D, colonies formed by MCF-7 cells with UCA1 overexpression and BT474 cells with UCA1 knockdown after treatment with 1 μ mol/L tamoxifen for 14 days (0.5% crystal violet staining, × 4); E, relative colony number and size of the MCF-7 and BT474 cell colonies in figure D; F, flow cytometry analysis of cell cycle distribution of MCF-7 cells and BT474 cells 72 h after treatment indicated in figure D; G and H, cell cycle distribution of MCF-7 and BT474 cells in figure F. * , compared with vector or miR-NC; #, compared with vector+Tam or si-NC+Tam; * and #, P< 0.05; * * and ##, P< 0.01. Tam, tamoxifen. |
MCF-7细胞在他莫昔芬处理后, miR-18a的表达水平显著下降(图3A), 在MCF-7细胞中过表达UCA1可以显著降低细胞中miR-18a的表达水平(图3B), 而在LCC2、LCC9和BT474细胞中, 敲减UCA1导致了4倍或以上的miR-18a表达增加(图3C)。生物信息学分析结果显示, UCA1可能有一个高度保守的miR-18a结合位点(图3D)。将野生型和突变型的UCA1片段插入pmirGLO荧光素酶表达载体后进行miR-18a结合实验结果显示, miR-18a可以显著抑制pmirGLO/UCA1-WT的荧光素酶表达, 但是并不能抑制pmirGLO/UCA1-MT的荧光素酶表达(图3E), 说明miR-18a可以结合预测的UCA1位点。
 | 图3 UCA1在乳腺癌细胞中竞争性结合miR-18aFigure 3 UCA1 competitively binds with miR-18aA, qRT-PCR analysis of miR-18a expression in MCF-7 cells after tamoxifen (0.1 μ mol/L) treatment at indicating time points up to 96 h; B, qRT-PCR analysis of miR-18a expression in MCF-7 cells 48 h after transection of UCA1 expression vector; C, qRT-PCR analysis of miR-18a expression in LCC2, LCC9, and BT474 cells 48 h after transfection of UCA1 siRNA or the negative control; D, putative binding site of miR-18a on lncRNA UCA1. The red nucleotides indicate the mutant sequence designed in the mutant lncRNA UCA1 constructs; E, the wildtype or the mutant UCA1 luciferase reporter constructs were co-transfected with miR-18a (100 nmol/L) or the scramble negative controls in MCF-7 cells. The relative firefly luciferase activity was measured 24 h after transfection and was normalized with renilla luciferase activity. * * P< 0.01, compared with the vector or the si-NC group. |
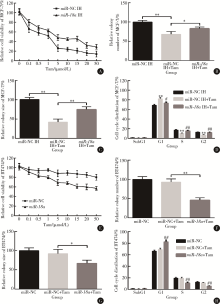
细胞活力和细胞克隆形成实验结果显示, 在MCF-7细胞中敲减miR-18a可以显著提高在不同浓度他莫昔芬处理后的细胞活力(图4A), 并且降低他莫昔芬对MCF-7细胞克隆数目及大小的抑制作用。相对于对照+他莫昔芬组, miR-18a抑制+他莫昔芬组的克隆数目多15.0%, 克隆平均大小增加33.0%(图4B、C)。敲减miR-18a也可以部分解除他莫昔芬诱导的G0/G1细胞周期阻滞。相对于对照+他莫昔芬组, miR-18a抑制+他莫昔芬组G1期细胞比例减少8.8%, S期细胞增加5.3%(图4D)。
 | 图4 miR-18a可以调控ER阳性乳腺癌细胞他莫昔芬敏感性Figure 4 miR-18a regulates sensitivity of ER-positive breast cancer cells to tamoxifenA and E, CCK-8 assay of cell viability of MCF-7 cells with or without miR-18a knockdown (A) and BT474 cells with or without miR-18a overexpression (E) after treatment with varying concentrations of tamoxifen (0.1, 0.5, 1.0, 5.0, 10.0, 15.0, 20.0, 50.0 μ mol/L) for 5 days; B, C, F, and G, quantification of relative colony number (B and F) and size (C and G) of MCF-7 cells with or without miR-18a knockdown (B and C) and BT474 cells with or without miR-18a overexpression (F and G) after treatment with 1 μ mol/L tamoxifen for 14 d; D and H, quantification of cell cycle distribution of MCF-7 cells with or without miR-18a knockdown (D) and BT474 cells with or without miR-18a overexpression (H) after treatment with 1 μ mol/L tamoxifen for 3 d. * , compared with miR-NC; #, compared with miR-NC+Tam; * and #, P< 0.05; * * and ##, P< 0.01. Tam, tamoxifen. |
在BT474细胞中转染miR-18a可以显著降低在不同浓度他莫昔芬处理后的细胞活力(图4E), 并且增加他莫昔芬对MCF-7细胞克隆数目及大小的抑制作用。相对于对照+他莫昔芬组, miR-18a过表达+他莫昔芬组的克隆数目少47.0%, 克隆平均大小减少25.0%(图4F、G)。过表达miR-18a可以提高他莫昔芬诱导的G0/G1细胞周期阻滞。相对于对照+他莫昔芬组, miR-18a过表达+他莫昔芬组G1期细胞比例增加13.3%, S期细胞减少7.9% (图4H)。
虽然ER阳性乳腺癌的内分泌治疗已经取得了很多进展, 但肿瘤对治疗产生的新的和获得性的耐药仍是重要的临床问题[20]。lncRNA对内分泌治疗的调控作用逐渐被人们所认知, 如BCAR4和HOTAIR的高表达均参与了乳腺癌细胞他莫昔芬耐药性的调控[10, 13]。以往的报道指出UCA1在乳腺癌中有明确的促癌作用[15, 17, 18], 并且UCA1也可以在多种肿瘤细胞中调控化疗敏感性, 如非小细胞肺癌细胞、乳腺癌细胞和肝癌细胞[19, 21, 22]。lncRNA的调控机制相当复杂, 且与下游分子参与的调控通路密切相关。
本研究发现, 他莫昔芬耐药的LCC2、LCC9和BT474细胞中的UCA1表达水平显著高于他莫昔芬敏感的MCF-7细胞。最近的另外一项研究也证实, 他莫昔芬耐药的乳腺癌细胞和其外泌体中的UCA1的表达水平上调[23]。功能上, UCA1过表达可以显著降低他莫昔芬敏感性, 而敲减UCA1则可以明显增加他莫昔芬敏感性。有研究指出, UCA1可以通过抑制mTOR信号途径增加他莫昔芬耐药性[24], 同时, 竞争性内源RNA也是lncRNA发挥其调控作用的重要机制。通过扮演分子海绵, lncRNA可以降低细胞内与其结合的miRNA水平, 并恢复miRNA抑制的信号通路[25]。在多种肿瘤细胞中, UCA1通过miRNA分子海绵作用调控了多种促癌通路, 如UCA1可以通过miR-145/ZEB1/2/FSCN1通路促进膀胱癌细胞迁移与侵袭[26], 也可以通过降低miR-193a-3p来促进非小细胞肺癌的发展[27], 还可以通过抑制miR-216b而激活FGFR1/ERK信号通路, 促进肝癌进展[28]。本研究发现, UCA1可以通过分子海绵作用降低miR-18a表达水平, 从而促进ER阳性乳腺癌细胞的他莫昔芬耐药性。
以前的研究报道, miR-18a在乳腺癌中起到肿瘤抑制分子作用。通过抑制HIF1-α 表达, miR-18a可以减少乳腺癌细胞肺转移[29], 更重要的是, miR-18a可以调控细胞周期进展。有报道指出, miR-18a可以抑制cdc42表达, 从而抑制肿瘤细胞的细胞周期进展, 减慢细胞增殖和侵袭[30], 因此, miR-18a很有可能是一个重要的UCA1下游细胞周期调控分子, 但是, 本文并未通过体外肿瘤模型对UCA1和miR-18a的他莫昔芬作用机制进行研究, 这是本研究的不足之处。UCA1在他莫昔芬处理后上调的机制还不清楚, 如是否受到ER信号途径调控也还未知, 除此以外, 由于lncRNA和miRNA均可以参与多信号途径调控, UCA1和miR-18a在乳腺癌中的调控网络仍需要更深入研究。
(本文编辑:任英慧)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|