

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
维生素D受体 FokⅠ多态性对牙周组织细胞CYP24A1表达的影响
[刘凯宁, 孟焕新△  , 侯建霞]
, 侯建霞]
, 侯建霞]
|
|


目的 维生素D受体(vitamin D receptor, VDR)基因的第二外显子存在唯一一个可以影响VDR蛋白结构的多态性位点,其可以由限制性核酸内切酶 FokⅠ所识别,分为FF、Ff和ff三型。 CYP24A1是维生素D 24羟化酶的编码基因,是常见的维生素D效应基因。本研究将探讨人牙龈成纤维细胞(human gingival fibroblasts,hGF)和牙周膜细胞(human periodontal ligament cells,hPDLC)中 VDR- FokⅠ多态性对CYP24A1表达的影响。方法 原代培养12名供体的hGF和hPDLC,提取基因组DNA,PCR扩增包含多态性位点的267 bp的片段。根据 FokⅠ对片段酶切的结果判断 VDR- FokⅠ基因型。确定基因型后,给各基因型hGF和hPDLC以10 nmol/L 1α,25双羟维生素D3(1,25OH2D3)或1 000 nmol/L 25羟维生素D3(25OHD3)刺激48 h,提取RNA,其中10 nmol/L 1,25OH2D3刺激48 h后还提取蛋白。之后给予hGF和hPDLC VDR拮抗剂ZK159222,再以10 nmol/L 1,25OH2D3或1 000 nmol/L 25OHD3刺激48 h,提取RNA。应用Real-time PCR和Western blot的方法检测维生素D24羟化酶CYP24A1和VDR的mRNA和蛋白的表达水平。结果 12名供体中,FF、ff和Ff型分别为4例、3例和5例。1,25OH2D3刺激hGF和hPDLC后,FF型细胞 CYP24A1的mRNA表达水平显著高于Ff型或ff型细胞(hGF: F=31.147, P<0.01;hPDLC: F=23.347, P<0.01);FF型细胞CYP24A1的蛋白表达水平同样显著高于Ff型或ff型细胞(hGF: F=12.368, P<0.01;hPDLC: F=15.749, P<0.01)。25OHD3刺激hGF和hPDLC后,FF型细胞 CYP24A1的mRNA表达水平也显著高于Ff型或ff型细胞(hGF: F=32.061, P<0.01;hPDLC: F=32.569, P<0.01)。如1,25OH2D3刺激伴有ZK159222,则三型细胞 CYP24A1的mRNA表达水平差异无统计学意义(hGF: F=0.246, P=0.787;hPDLC: F=0.574, P=0.583)。如25OHD3刺激伴有ZK159222,则三型细胞 CYP24A1的mRNA表达水平差异也无统计学意义(hGF: F=1.636, P=0.248;hPDLC: F=0.582, P=0.578)。不同刺激条件下,hGF和hPDLC两种细胞比较CYP24A1或VDR的表达水平,差异均无统计学意义。结论 在hGF和hPDLC中,FF型VDR可介导比其他基因型VDR更为显著的CYP24A1上调,提示FF型VDR可能具有更强的转录活性。
Objective: There is asingle nucleotide polymorphism (SNP) in the exon 2 of the vitamin D receptor ( VDR) gene that can be distinguished using the restriction endonuclease FokⅠ, and accordingly divided into three genotypes: FF, Ff and ff. VDR- FokⅠ polymorphism was the only known SNP that could alter the protein structure of VDR. CYP24A1 is the gene encoding vitamin D 24 hydroxylase and is a vitamin D responsive gene. The influence of rs2228570 on transcriptional activation by VDR in human gingival fibroblasts (hGF) and periodontal ligament cells (hPDLC) was investigated in this study.Methods: hGF and hPDLC of 12 donors’ were primarily cultured and genomic DNA was extracted. A part of genomic DNA with the length of 267 bp was obtained using PCR, which contained the SNP. VDR- Fok Ⅰ genotypes were determined according to the results of restriction fragment length polymorphism. hGF and hPDLC were stimulated with 10 nmol/L 1α,25 dihydroxy vitamin D3 (1,25OH2D3) or 1 000 nmol/L 25 hydroxy vitamin D3 (25OHD3) for 48 h before RNA was extracted. Then VDR antagonist ZK159222 was used or not used during 1,25OH2D3 or 25OHD3 stimulation with hGF and hPDLC. After 1,25OH2D3 stimulation for 48 h, the proteins in hGF and hPDLC were also collected. The protein expressions of CYP24A1 and VDR were detected using Western blot.Results: Among the 12 donors’ cell cultures, the number of FF, ff and Ff genotypes was 4, 3 and 5, respectively.After stimulation with 1,25OH2D3or 25OHD3 for 48 h, CYP24A1 mRNA levels in FF-hGF were significantly higher than those in other hGF genotypes(1,25OH2D3: F=31.147, P<0.01; 25OHD3: F = 32.061, P <0.01), as was in FF-hPDLC (1,25OH2D3: F=23.347, P<0.01; 25OHD3: F = 32.569, P < 0.01). When ZK159222 was used before 1,25OH2D3 stimulation, this statistically significant difference disappeared (hGF: F=0.246, P=0.787; hPDLC: F=0.574, P=0.583). When ZK159222 was used before 25OHD3 stimulation, the trend was similar (hGF: F = 1.636, P = 0.248; hPDLC: F =0.582, P=0.578).After stimulation with 1,25OH2D3 for 48 h, CYP24A1 protein levels in FF-hGF were significantly higher than those in the other hGF genotypes ( F=12.368, P <0.01), as was in FF-hPDLC ( F=15.749, P <0.01). In hGF and hPDLC, the mRNA or protein expression of VDR of different genotypes was not significantly different under different stimulation conditions.The paired comparison showed that there was no statistically significant difference between the expression of CYP24A1 in hGF and that in hPDLC under all the stimulation conditions, as was the expression of VDR.Conclusion: In hGF and hPDLC, the FF-VDR genotype is associated with the more remarkable up-regulation of CYP24A1than the other genotypes, indicating that transcriptional activation of FF-VDR might be higher than those of other vitamin D receptors.
25羟维生素D3(25OHD3)是人体中维生素D3的主要存在形式, 其由1, α 羟化酶CYP27B1活化形成1α , 25双羟维生素D3(1, 25OH2D3)这一人体中维生素D3的活性形式, 在钙磷代谢和免疫功能调控中发挥重要作用[1, 2]。1, 25OH2D3通过维生素D受体(vitamin D receptor, VDR)发挥生物学作用, VDR包含N末端的DNA结合域和C末端的蛋白结合域[3]。1, 25OH2D3与VDR结合后, 首先与类视黄醇X受体形成异二聚体, 并进一步募集转录因子Ⅱ B发挥稳定RNA聚合酶Ⅱ 前复合体的作用, 从而促成维生素D靶基因的转录[4]。
VDR基因第二外显子起始密码子上游第8个碱基对处存在T/C多态性位点(rs2228570), 能够为限制性内切酶FokⅠ 所识别, 如果能被FokⅠ 切开记为F, 不能切开记为f。F型和f型分别对应424个氨基酸的短VDR蛋白和427个氨基酸的长VDR蛋白。VDR-FokⅠ 多态性位点是已知的VDR基因中唯一能造成蛋白结构改变的单核苷酸多态性位点, 且与已知其他位点没有连锁关系[5, 6], 其对VDR功能的影响值得深入研究。Arai等[7]最先通过同时转染不同VDR-FokⅠ 的表达质粒的方法, 发现F型VDR的转录活性约是f型VDR转录活性的1.7倍。其后陆续有研究利用类似的方法获得了F-VDR转录活性强于f-VDR的结果[4, 8, 9]。然而也有研究利用同样的方法却并未发现F-VDR与f-VDR转录活性存在差异[10, 11], 因此, VDR-FokⅠ 多态性对VDR转录活性的影响尚不明确, 需要进一步研究。
目前已有研究发现VDR-FokⅠ 多态性与侵袭性牙周炎相关。韩国学者Park等[12]纳入了93名广泛性侵袭性牙周炎患者和143名健康对照, 国内学者Li等[13]纳入了51名广泛性侵袭性牙周炎患者和53名健康对照, 研究均发现FF型与广泛性侵袭性牙周炎正相关, 但目前未见探索VDR-FokⅠ 多态性与侵袭性牙周炎相关性机制的研究。本课题组之前的研究纳入了66名广泛性侵袭性牙周炎患者、52名慢性牙周炎患者、60名健康对照, 同样发现FF基因型与广泛性侵袭性牙周炎正相关; 进一步的研究发现FF型的人牙周膜细胞(human periodontal ligament cells, hPDLC)受1, 25OH2D3刺激后碱性磷酸酶和骨钙素表达显著高于Ff型和ff型[14], 这一研究证实VDR-FokⅠ 多态性能够影响牙周组织细胞矿化相关因子的表达, 但仍不能直接解释VDR-FokⅠ 多态性与侵袭性牙周炎相关的机制。
维生素D24羟化酶可以使1, 25OH2D3的24位碳原子加羟基从而活性大大降低, 是人体中1, 25OH2D3活性的抑制酶, 对1, 25OH2D3的生物学效应进行负向调控, 其编码基因为CYP24A1[3]。CYP24A1是常见的维生素D效应基因, 其启动子区具有维生素D反应元件, 在1, 25OH2D3存在时显著上调[1]。本研究将在此基础上进一步探索两种人牙周组织细胞人牙龈成纤维细胞(human gingival fibroblasts, hGF)和hPDLC中VDR-FokⅠ 多态性对CYP24A1表达的影响, 进一步分析VDR-FokⅠ 多态性对VDR转录活性的影响, 以期为VDR-FokⅠ 多态性与侵袭性牙周炎相关性的机制研究提供科学线索。
本研究经北京大学口腔医院生物医学伦理委员会同意, 伦理审查批准号为PKUSSIRB-2011007, 所有志愿者均知晓研究目的并签署知情同意书。
普通杜尔伯科改良伊格尔培养基(Dulbecco’ s modified Eagle’ s medium, DMEM)购于美国Gibco公司, 无酚红DMEM购于美国Sigma公司, 普通胎牛血清(fetal bovine serum, FBS)购于美国Hyclone公司, 活性炭处理的FBS购于中国灏洋生物公司, FokⅠ 限制性内切酶购于日本TAKARA公司, 25OHD3和1, 25OH2D3购于美国Sigma公司, Trizol购于美国Invitrogen公司, SYBR绿色荧光染料(SYBR Green)实时定量PCR(real-time PCR, RT-PCR)试剂盒购于瑞士Roche公司, CYP24A1单克隆抗体和VDR多克隆抗体购于美国Santa Cruz公司, VDR拮抗剂ZK159222获赠于德国Bayer公司Ekkehard May博士及其同事。
牙龈组织及牙周膜组织来源于12名健康志愿者, 参照本课题组以往的方法[15, 16], 进行hGF和hPDLC原代培养。培养条件为含10%(体积分数)FBS的DMEM培养基, 加入100 U/mL 青霉素和100 mg/L 链霉素, 在5%(体积分数)CO2、37 ℃恒温培养箱中培养。待原代细胞长至80%时, 用 0.25%(质量分数)胰蛋白酶和0.02%(质量分数)乙二胺四乙酸消化, 1 ∶ 2比例传代, 4~6代细胞用于后续实验。
提取hGF和hPDLC的基因组DNA, PCR扩增VDR片段, 上游引物为5'-AGCTGGCCCTGGCACTGACTCTGGCTCT-3', 下游引物为5'-ATGGAAACACCTTGCTTCTTCTCCCTC-3'。反应条件:95 ℃预变性10 min; 95 ℃变性30 s, 65 ℃退火30 s, 72 ℃延伸30 s, 共35个循环; 最后72 ℃延伸7 min。PCR产物为267 bp。
应用限制性内切酶Fok Ⅰ 对PCR产物进行消化, 在267 bp的PCR产物中有1个固定的Fok Ⅰ 酶切位点, 可将产物分作195 bp和72 bp两段, F表示PCR产物在该位点不能被Fok Ⅰ 切开, f表示PCR产物在该位点能被Fok Ⅰ 切开, 因此, 3种基因型的消化产物分别为:FF基因型只有267 bp一种产物; ff基因型有195 bp和72 bp两种产物; 杂合子Ff基因型有267 bp、195 bp和72 bp三种产物。酶切产物经2.5%(质量分数)琼脂糖凝胶电泳, 溴乙锭染色后观察结果。
以下研究中培养hGF和hPDLC的培养基均为含10%(体积分数)活性炭处理FBS的无酚红DMEM, 共分为5组:
组1首先给予0.1%(体积分数)乙醇1 h, 换液后给予细胞10 nmol/L 1, 25OH2D3刺激, 48 h后刺激终止。
组2首先给予细胞1 000 nmol/L VDR拮抗剂ZK159222[17, 18]刺激1 h, 换液后给予细胞10 nmol/L 1, 25OH2D3刺激, 48 h后刺激终止。
组3首先给予0.1%(体积分数)乙醇1 h, 换液后给予细胞1 000 nmol/L 25OHD3刺激, 48 h后刺激终止。
组4首先给予细胞1 000 nmol/L VDR拮抗剂ZK159222刺激1 h, 换液后给予细胞1 000 nmol/L 25OHD3刺激, 48 h后刺激终止。
组5为对照组, 给予细胞0.1%(体积分数)乙醇49 h。
细胞刺激终止后, Trizol提取RNA。逆转录试剂盒合成cDNA, 保存于-20 ℃待用。使用SYBR Green试剂盒进行RT-PCR实验, 检测CYP24A1和VDR的mRNA表达, 以GAPDH为内参。mRNA表达水平应用公式2-Δ Δ Ct表示, 所用引物见表1。
| 表1 实时定量PCR引物序列 Table 1 List of primer sequences used for real-time PCR |
1, 25OH2D3刺激组的细胞刺激终止后, hGF和hPDLC两种细胞中每基因型取3例进行细胞裂解提取蛋白, 应用BCA试剂盒测定蛋白浓度后, 每样本取20 μ g用于Western blot实验。检测CYP24A1的一抗为小鼠来源的单克隆抗体, 稀释比例为1 ∶ 1 000; 检测VDR的一抗为兔来源的多克隆抗体, 稀释比例为1 ∶ 1 000; 以β -actin为内参, 其检测一抗为小鼠来源的单克隆抗体, 稀释比例为1 ∶ 1 000; 检测小鼠或兔蛋白的二抗稀释比例为1 ∶ 2 500。
采用单因素方差分析比较不同基因型细胞CYP24A1及VDR的mRNA、蛋白表达水平, 并用LSD法进行两两比较。用配对t检验法比较hGF和hPDLC两种细胞的CYP24A1和VDR的mRNA、蛋白表达水平。统计学分析使用SPSS 11.5软件完成, P< 0.05认为差异有统计学意义。
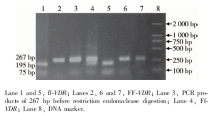
12名供体的细胞中, FF、Ff和ff基因型分别有4例、5例和3例。图1所示为PCR结果, 第1~6泳道均为PCR产物; 第7泳道为PCR的阴性对照, 说明无污染; 第8泳道为DNA marker。图2所示为FokⅠ 酶切结果, 第1、5泳道为ff型; 第2、6、7泳道为FF型; 第3泳道为未经酶切的PCR产物; 第4泳道为Ff型; 第8泳道为DNA marker。
 | 图1 267 bp的PCR产物的琼脂糖凝胶电泳结果Figure 1 Agarose gel electrophoresis of PCR products of 267 bp |
 | 图2 不同VDR-FokⅠ 基因型的琼脂糖凝胶电泳结果Figure 2 Agarose gel electrophoresis of different genotypes of VDR |
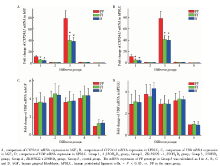
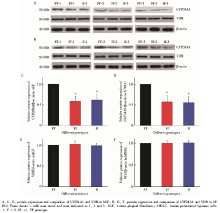
图3中, 在FF型hGF(图3A, 组1, F=31.147, P< 0.01)和FF型hPDLC(图3B, 组1, F=23.347, P< 0.01)受1, 25OH2D3刺激后, CYP24A1的mRNA表达水平均高于Ff型和ff型同种细胞; CYP24A1的蛋白表达水平也均高于Ff型和ff型同种细胞(图4, hGF:图4A, C, F=12.368, P< 0.01; hPDLC:图4B, D, F=15.749, P< 0.01)。在应用VDR拮抗剂ZK159222后, 再用1, 25OH2D3刺激hGF(图3A, 组2, F=0.246, P=0.787)和hPDLC(图3B, 组2, F=0.574, P=0.583)时, 不同VDR-FokⅠ 基因型细胞CYP24A1的mRNA表达水平差异无统计学意义。
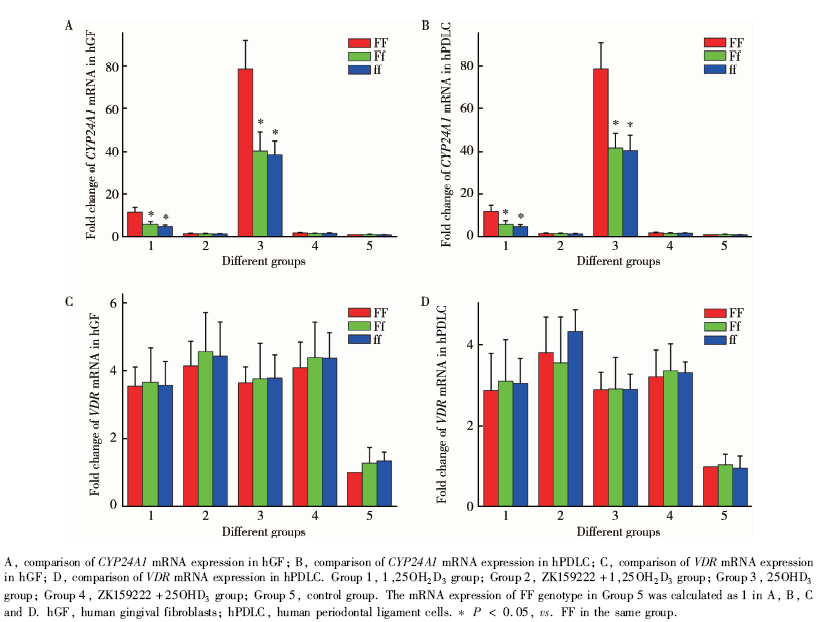 | 图3 VDR-FokⅠ 基因型对hGF和hPDLC中CYP24A1和VDR的mRNA表达水平的影响Figure 3 Influence of VDR-FokⅠ polymorphism on the mRNA expression of CYP24A1 and VDR in hGF and hPDLC |
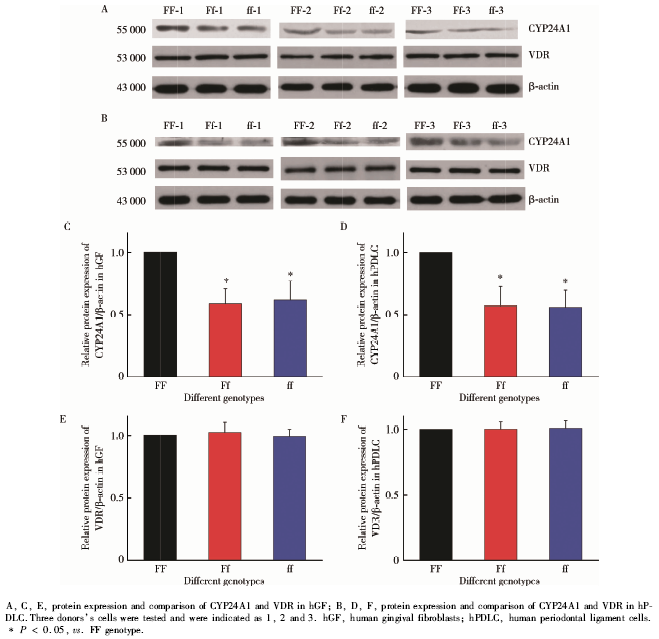 | 图4 VDR-FokⅠ 基因型对hGF和hPDLC中CYP24A1和VDR的蛋白表达水平的影响Figure 4 Influence of VDR-FokⅠ polymorphism on the protein expression of CYP24A1 and VDR in hGF and hPDLC |
给予hGF(图3A, 组3, F=32.061, P< 0.01)和hPDLC(图3B, 组3, F=32.569, P< 0.01)25OHD3刺激, FF型细胞CYP24A1的mRNA表达水平高于Ff型和ff型细胞; 在应用VDR拮抗剂ZK159222后, 再用25OHD3刺激hGF(图3A, 组4, F=1.636, P=0.248)和hPDLC(图3B, 组4, F=0.582, P=0.578)时, 不同VDR-FokⅠ 基因型细胞CYP24A1的mRNA表达水平差异无统计学意义。
在所有组别中, 不同VDR-FokⅠ 基因型细胞VDR的mRNA及蛋白表达水平差异均无统计学意义(图3C、3D、4E、4F)。
如图5所示, 对所有组别中hGF和hPDLC中CYP24A1的mRNA和蛋白表达水平(图5A、C)进行两种细胞间的两两比较, 差异均无统计学意义; 对各组hGF和hPDLC中VDR的mRNA和蛋白表达水平(图5B、D)进行两两比较, 差异也均无统计学意义。
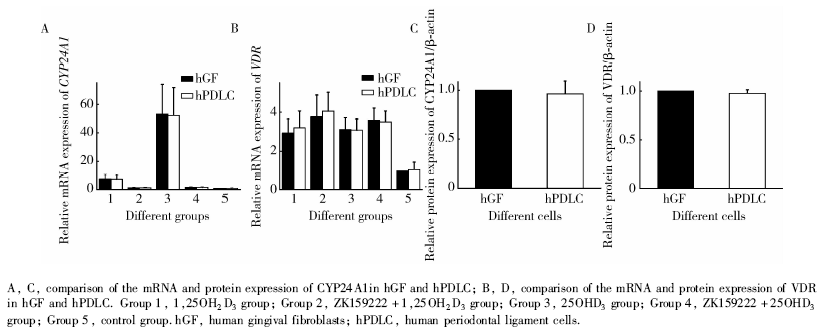 | 图5 hGF和hPDLC中CYP24A1和VDR的mRNA和蛋白表达水平的比较Figure 5 Comparison of the mRNA and protein expression of CYP24A1 and VDR in hGF and hPDLC |
因为CYP24A1是常见的维生素D效应基因, 本课题组既往研究已经证实hGF和hPDLC中CYP24A1在1, 25OH2D3刺激时可显著上调[15], 所以本研究选择CYP24A1作为检验VDR转录活性的工具。同时本课题组的既往研究[15]分别应用1 000 nmol/L的25OHD3和10 nmol/L的1, 25OH2D3刺激hGF和hPDLC, 即发现1 000 nmol/L的25OHD3促进CYP24A1的mRNA表达水平上调的作用显著强于10 nmol/L的1, 25OH2D3, 与本文的结果类似。分析其原因, 可能是组1中的1, 25OH2D3刺激需进入细胞后才能发挥生物学作用, 而组3中25OHD3刺激后hGF和hPDLC产出的1, 25OH2D3在细胞内即可与VDR结合发挥生物学作用, 作用更为直接所以作用更强。
本研究发现, 1, 25OH2D3或25OHD3刺激后, FF型hGF和hPDLC表达CYP24A1的水平显著高于Ff型和ff型细胞, 而应用VDR拮抗剂阻断VDR功能, 均使得3种基因型细胞CYP24A1的表达水平差异无统计学意义, 这提示FF型VDR可能较另外两型VDR具有更高的转录活性。与此同时, 我们还比较了多种刺激条件下3种基因型细胞VDR mRNA和蛋白的表达水平, 差异均无统计学意义。由此可见, FF型hGF和hPDLC表达CYP24A1的水平显著高于Ff型和ff型细胞的原因不是VDR表达水平不同, 从而从另一个侧面提示FF型VDR转录活性可能高于Ff型和ff型VDR。ff型VDR比FF型VDR多3个氨基酸, 其中多出的第2个氨基酸为谷氨酸, 其所带的负电荷可能妨碍VDR的N末端和转录因子Ⅱ B的结合, 从而使得ff型VDR提供转录因子Ⅱ B 参与mRNA合成的能力弱于FF型VDR[4]。这一分子机制可能是FF型VDR具有更高转录活性的原因。
之前有少量VDR-FokⅠ 基因型对VDR功能影响的研究, 但其中多数研究使用了外源性VDR表达质粒[4, 7, 8, 9, 11]。而Whitfield等[19]的研究发现, 应用外源性VDR和细胞内源性VDR进行转录活性研究, 结果并不相同, 提示转染外源性VDR表达质粒的研究结果和体内的实际情况可能存在一定差异, 因此, 本研究选择细胞内源性的不同FokⅠ 基因型的VDR作为研究对象进行转录活性的探索, 以期更接近于体内的实际情况, 但不同供体来源的细胞中除VDR-FokⅠ 多态性不同外, 可能还存在其他未知的影响, 也非完美无缺的研究模型, 所以需要更多的研究进行进一步的探索。
VDR-FokⅠ 基因多态性可产生FF、Ff和ff 3种基因型。杂合子Ff型可以同时产生427个氨基酸的长VDR蛋白和424个氨基酸的短VDR蛋白。在多数既往的研究中, 只涉及纯合子FF型和ff型的比较[4, 8, 9, 11]。只有Arai等[7]的研究中同时转染FF型和ff型的VDR表达质粒以模拟Ff型, 发现Ff型VDR的转录活性显著高于ff型并显著低于FF型。本研究发现Ff型细胞受1, 25OH2D3刺激后CYP24A1表达显著低于FF型, 而与ff型差异无统计学意义, 因为本研究对象为细胞内源性VDR, 所以我们的结果可能比Arai等[7]的研究结果更接近于体内实际的情况。
本研究中我们使用的hGF和hPDLC来自于相同的12名志愿者, 进行两种细胞之间的比较时, 可以避免不同供体来源的影响。在本课题组之前的研究中[14], hGF在受1, 25OH2D3刺激后, 碱性磷酸酶和骨钙素的表达均显著低于hPDLC。究其原因, 可能还是因为hPDLC比hGF更接近成骨细胞的特点[20], 所以hPDLC的矿化相关因子表达更高。本研究发现两种细胞的CYP24A1表达差异无统计学意义, 因此两种细胞中VDR的转录活性差异可能并无统计学意义。
除了与侵袭性牙周炎相关, VDR-FokⅠ 基因多态性还被发现与严重呼吸道合胞病毒性细支气管炎[21]、酗酒[22]等相关, 然而由于缺乏功能研究, 具体机制均不清楚。本研究发现FF型hGF和hPDLC受1, 25OH2D3刺激时, CYP24A1上调更为显著, 提示FF型VDR可能具有更强的转录活性, 该发现为解释VDR-FokⅠ 基因多态性与侵袭性牙周炎的相关性提供了线索。研究者下一步将进行更多的功能研究, 以便进一步深入理解VDR-FokⅠ 基因多态性与侵袭性牙周炎的关系。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|