
1 资料与方法
1.1 病例选择和标本
1.2 组织芯片制备和免疫组织化学染色
1.3 构建LASS2全长或突变质粒
表1 构建质粒所用引物序列Table 1 Nucleotide sequences of primers used in plasmid construct |
| Gene | Primer |
| LASS2 | Forward: 5′-GGGGACAAGTTTGTACAAAAAAGCAGGCTTAATGCTCCAGACCTTGTATGATTA-3′ |
| Reverse: 5′-GGGGACCACTTTGTACAAGAAAGCTGGGTAGTCATTCTTACGATGGTTGT-3′ | |
| LASS2-T332A | Forward: 5′-TACCAGCTTTCCAGCTATGAACTTGTGGGCCATG-3′ |
| Reverse: 5′-CATGGCCCACAAGTTCATAGCTGGAAAGCTGGTA-3′ | |
| LASS2-S341A | Forward: 5′-TCTGTTTCTTCCCGGTCAGCGCGTTCATCTTCTACCAG-3′ |
| Reverse: 5′-CTGGTAGAAGATGAACGCGCTGACCGGGAAGAAACAGA-3′ | |
| LASS2-T346A | Forward: 5′-CTCTGAGCTCTCTGCTTCTTCCCGGTCACTGC-3′ |
| Reverse: 5′-GCAGTGACCGGGAAGAAGCAGAGAGCTCAGAG-3′ | |
| LASS2-S348A | Forward: 5′-TCCTCCCCCTCTGAGGCCTCTGTTTCTTCCCG-3′ |
| Reverse: 5′-CGGGAAGAAACAGAGGCCTCAGAGGGGGAGGA-3′ | |
| LASS2-S349A | Forward: 5′-CCTCCTCCCCCTCTGCGCTCTCTGTTTCTTC-3′ |
| Reverse: 5′-GAAGAAACAGAGAGCGCAGAGGGGGAGGAGG-3′ |
LASS2, homo sapiens longevity assurance homolog 2 of yeast LAG1. |
1.4 质谱法分析LASS2蛋白的磷酸化位点
1.5 细胞培养和转染
1.6 细胞生长曲线
1.7 MTT掺入实验
1.8 平板克隆形成实验
1.9 细胞划痕修复试验
1.10 Transwell细胞侵袭实验
1.11 细胞周期检测
1.12 细胞凋亡检测
1.13 Western blot实验
1.14 免疫共沉淀实验
1.15 谷胱甘肽-S-转移酶融合蛋白沉降技术(GST pull-down)
1.16 V-ATPase活性的检测
1.17 细胞外H+浓度测定
1.18 明胶酶谱实验
1.19 统计学分析方法
2 结果
2.1 LASS2是前列腺癌的潜在预后标记物
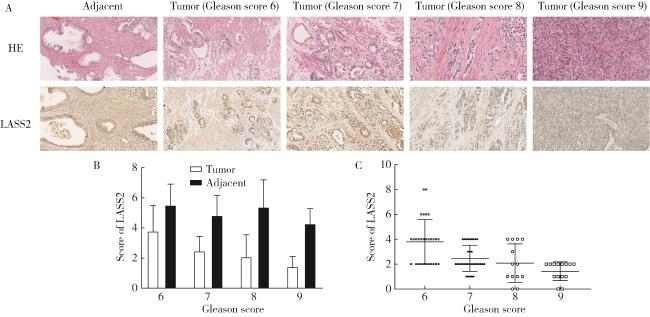
图1 LASS2在前列腺癌及癌旁组织中的表达Figure 1 Expression level of LASS2 in prostate cancer and adjacent tissue A, immunohistochemical staining of LASS2 in 90 prostate carcinoma samples with paired adjacent normal tissues. Representative sections from prostate cancer (Gleason score 6, 7, 8, 9) or adjacent normal tissue stained with HE and LASS2 antibody are presented (×200). B, LASS2 levels was significantly downregulated in prostate cancer tissues compared with paired adjacent normal tissues (P < 0.01); C, LASS2 levels decreased with increasing Gleason scores of prostate cancer tissues (P < 0.01). HE, hematoxylin-eosin; LASS2, homo sapiens longevity assurance homolog 2 of yeast LAG1. |
2.2 LASS2的C末端存在3个丝氨酸磷酸化位点
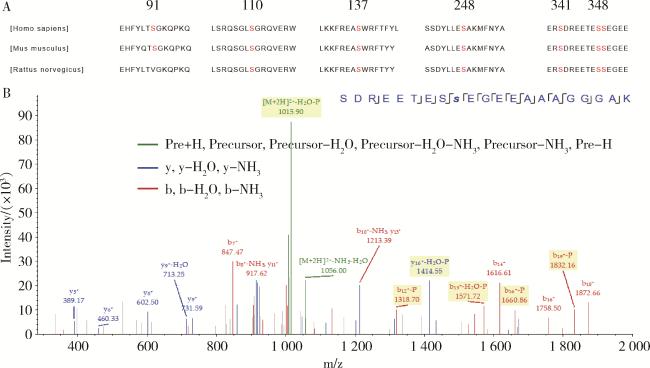
图2 LASS2的C末端丝氨酸磷酸化位点Figure 2 Serine phosphorylation site at the C-terminus of LASS2 A, protein analysis using PROSITE (http://prosite.expasy.org/) predicted that LASS2 contained seven protein serine phosphorylation sites, and these phosphorylation sites are highly conserved among human, mouse and rat; B, FLAG-LASS2 was purified from transfected HEK 293T cells in the presence of phosphatase inhibitor and subjected to mass spectrometric analysis. Phosphoproteomic study has identified three serine phosphorylation sites in the C-terminal of LASS2. LASS2, homo sapiens longevity assurance homolog 2 of yeast LAG1. |
2.3 LASS2-S348A去磷酸化突变体促进PC-3M-1E8细胞的生长、迁移和侵袭
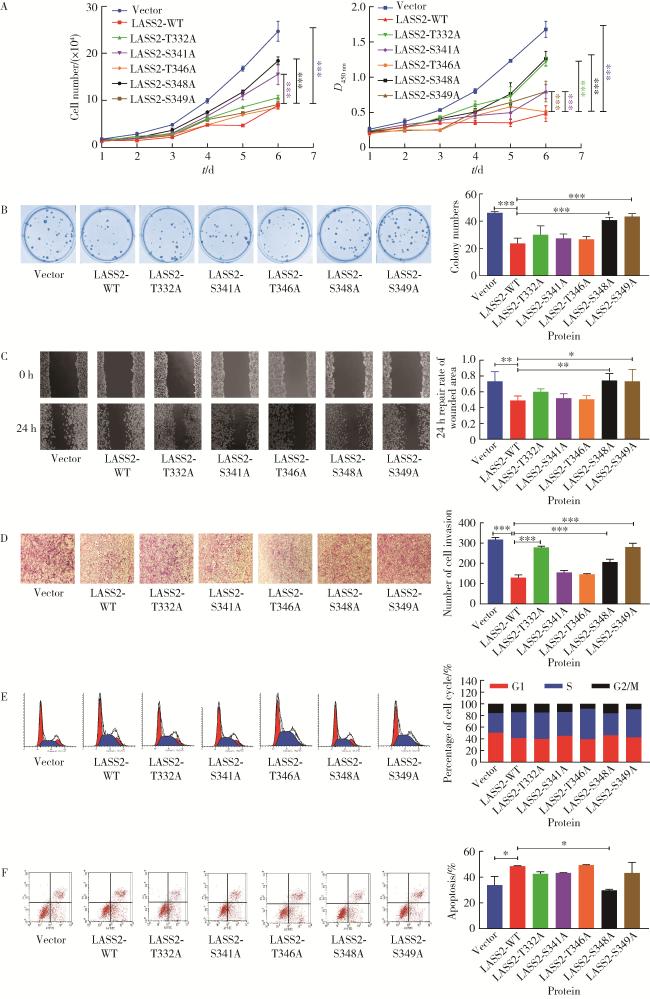
图3 LASS2-WT及其突变体对前列腺癌细胞系PC-3M-1E8生长、迁移和侵袭能力的影响Figure 3 Effects of LASS2-WT and its mutants on the growth, migration, and invasion ability of prostate cancer cell line PC-3M-1E8 A, growth curve assay and MTT test (n=3); B, cell cloning experiment (n=3); C, cell migration experiment (n=4); D, cell invasion experiment (n=3); E, cell cycle experiment; F, cell apoptosis experiment (n=3). These figures showed dephosphorylation of LASS2 at serine residue 348 promote growth, migration and invasion of PC-3M-1E8 cells. LASS2, homo sapiens longevity assurance homolog 2 of yeast LAG1; *P < 0.05, * *P < 0.01, * * *P < 0.001 vs. LASS2-WT. |
2.4 LASS2 S348A去磷酸化突变体增强V-ATPase活性和细胞外H+浓度
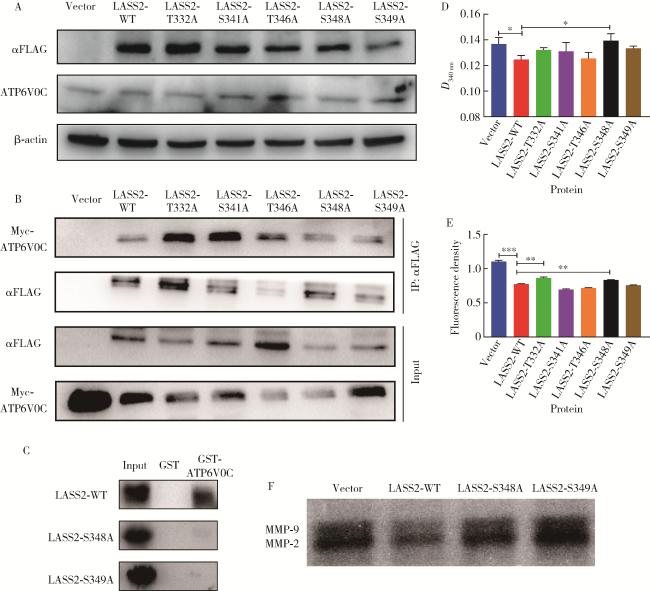
图4 LASS2-WT及其突变体对前列腺癌细胞系PC-3M-1E8 V-ATPase活性和细胞外H+浓度的影响Figure 4 Effects of LASS2-WT and its mutants on V-ATPase activity and extracellular H+ concentration in prostate cancer cell line PC-3M-1E8 A, Western blotting; B, co-immunoprecipitation; C, GST pull-down; D, V-ATPase activity detection experiment (n=3); E, extracellular H+ detection experiment (n=6); F, gelatin enzyme spectrum experiment. These figures showed that LASS2-S348A dephosphorylation mutant enhances V-ATPase activity and extracellular H+ concentration. LASS2, homo sapiens longevity assurance homolog 2 of yeast LAG1; WT, wild type; V-ATPase, vacuolar ATPase; ATP6V0C, C subunit of V0 domain of the V-ATPase; GST, glutathione-S-transferase; MMP, matrix metalloproteinase. *P < 0.05, * *P < 0.01, * * *P < 0.001. |
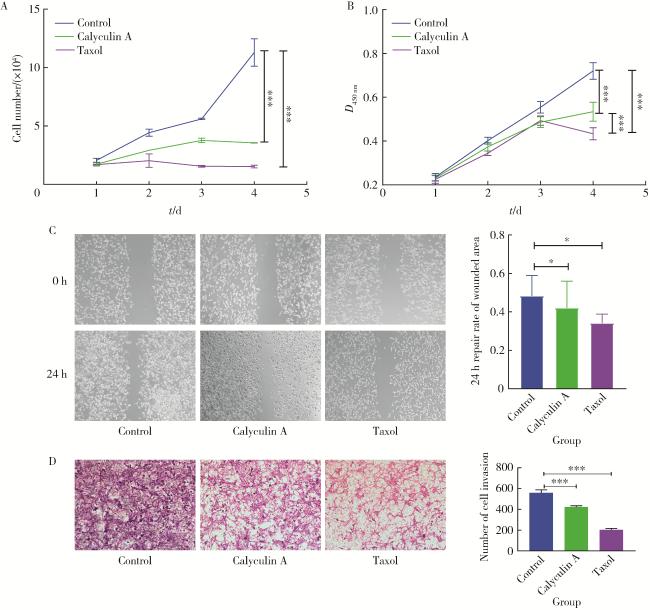
2.5 蛋白磷酸酶抑制剂calyculin A明显抑制前列腺癌细胞的增殖、迁移和侵袭
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
图5 Calyculin A或紫杉醇对前列腺癌细胞系PC-3M-1E8细胞增殖、迁移和侵袭的影响Figure 5 Effects of calyculin A or taxol on proliferation, migration, and invasion of prostate cancer cell line PC-3M-1E8 A, growth curve assay; B, MTT test; C, cell migration experiment (n=8); D, cell invasion experiment (n=3). Dimethyl sulfoxide (DMSO) was the negative control. These figures showed that calyculin A significantly inhibits the proliferation, migration, and invasion of PC-3M-1E8 cells. *P < 0.05, * * *P < 0.001, vs. LASS2-WT. |