
1 资料与方法
1.1 scRNA-seq数据集的获取
1.2 单细胞测序数据的质量控制及预处理
1.3 B细胞亚群注释
1.4 差异基因聚类分析
1.5 富集分析
1.6 PPI网络构建
1.7 统计学分析
2 结果
2.1 SS患者和健康对照外周血B细胞图谱构建
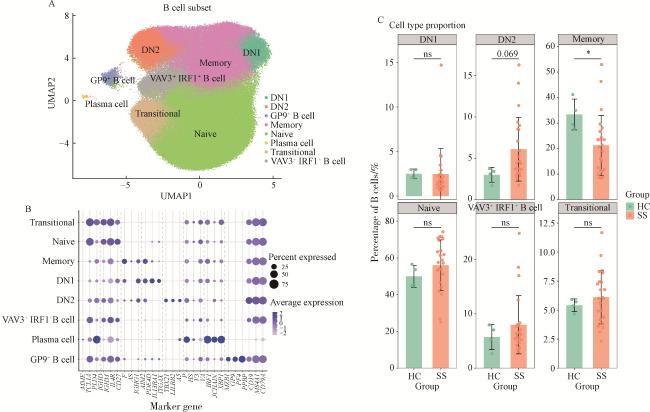
图1 SS患者和健康对照的B细胞亚群的分子鉴定及可视化分析Figure 1 Identification and molecular characterization of B cell subsets of SS patients and HC A, UMAP plots of B cell subsets; B, dot plot showing marker gene expression across B cell subsets; C, comparative analysis of B cell subsets between SS patients and HC. *P≤0.05; ns, no significant, P>0.05. SS, Sjögren syndrome; HC, healthy control; UMAP, uniform manifold approximation and projection; DN1, double negative 1 B cell; DN2, double negative 2 B cell; Memory, memory B cell; Naive, naïve B cell; Transitional, transitional B cell. |
2.2 基于无监督层次聚类的SS患者分子分型构建及富集分析
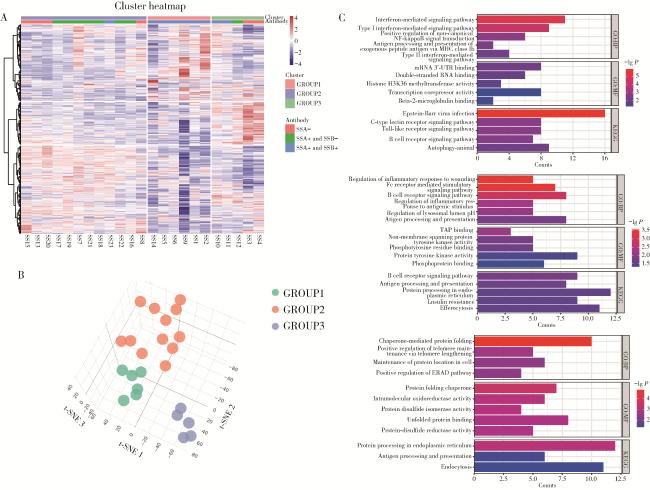
图2 基于B细胞基因表达谱的患者分型及其功能富集分析Figure 2 Molecular subtyping and enrichment analysis of patients based on differentially expressed genes A, integrated visualization from unsupervised consensus clustering on DEGs: molecular subtypes and anti-SSA/anti-SSB antibody status in SS patients; B, 3D t-SNE visualization of transcriptional stratification in SS subtypes; C, enrichment analysis across SS molecular subtypes via GO analysis (including biological processes and molecular functions) and KEGG pathway analysis. SS, Sjögren syndrome; SSA, Sjögren syndrome antigen A; SSB, Sjögren syndrome antigen B; DEGs, differentially expressed genes; t-SNE, t-distributed stochastic neighbor embedding; GO, Gene Ontology; BP, Biological Processes; MF, Molecular Functions; KEGG, Kyoto Encyclopedia of Genes and Genomes; GROUP1, interferon-dominant subtype; GROUP2, B cell activation subtype; GROUP3, endoplasmic reticulum stress subtype; Counts, the number of DEGs enriched in each GO or KEGG term. |
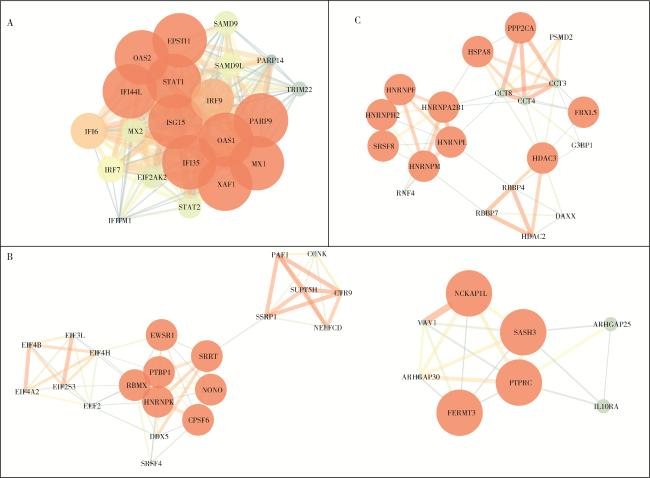
2.3 各亚型特征基因集的PPI网络构建及富集分析
2.4 不同亚型SS患者自身抗体及B细胞亚群特征
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
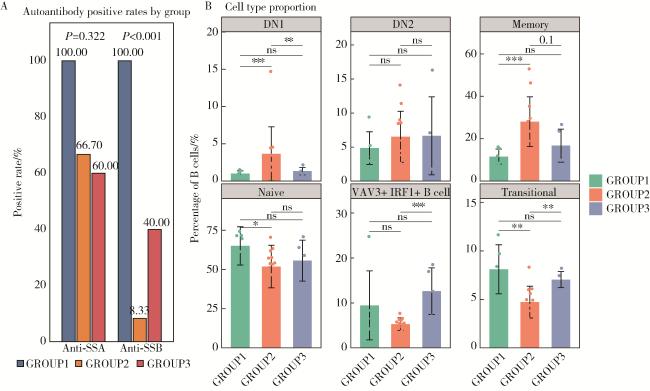
图4 SS不同分子亚型患者血清学自身抗体及外周血B细胞亚群特征比较Figure 4 Comparison of serological autoantibodies and peripheral blood B cell subset characteristics among different subtypes of SS patients A, subtype-specific positivity rates of anti-SSA and anti-SSB antibodies; B, proportions of peripheral blood B cell subsets within total B cells in SS subtypes. *P≤0.05; * *P≤0.01; * * *P≤0.001; ns, no significant, P>0.05. GROUP1, interferon-dominant subtype; GROUP2, B cell activation subtype; GROUP3, endoplasmic reticulum stress subtype; DN1, double negative 1 B cell; DN2, double negative 2 B cell; Memory, memory B cell; Naive, naïve B cell; Transitional, transitional B cell; SS, Sjögren syndrome; SSA, Sjögren syndrome antigen A; SSB, Sjögren syndrome antigen B; HC, healthy control. |
表1 三组患者抗SSA抗体与抗SSB抗体阳性率比较Table 1 Comparison of positive rates of anti-SSA and anti-SSB antibodies in three groups of patients |
| Items | Anti-SSA antibodies positive rate (positive/total) | Anti-SSB antibodies positive rate (positive/total) |
| GROUP1 | 100.00% (6/6) | 100.00% (6/6) |
| GROUP2 | 66.67% (8/12) | 8.33% (1/12) |
| GROUP3 | 60.00% (3/5) | 40.00% (2/5) |
| P value | 0.322 | < 0.001 |
GROUP1, interferon-dominant subtype; GROUP2, B cell activation subtype; GROUP3, endoplasmic reticulum stress subtype; SSA, Sjögren syndrome antigen A; SSB, Sjögren syndrome antigen B. |
表2 不同亚型SS患者B细胞亚群分布特征Table 2 Distribution characteristics of B cell subsets in different subtypes of SS patients |
| B cell subset | GROUP1 | GROUP2 | GROUP3 | GROUP1 vs. GROUP2 | GROUP1 vs. GROUP3 | GROUP2 vs. GROUP3 |
| DN1/%, $\bar x \pm s$ | 1.01±0.38 | 3.66±3.63 | 1.33±0.49 | < 0.001 | 0.329 | 0.004 |
| DN2/%, $\bar x \pm s$ | 4.85±2.39 | 6.51±3.74 | 6.65±5.72 | 0.553 | 0.792 | 0.879 |
| Memory/%, $\bar x \pm s$ | 11.6±3.51 | 28.01±11.75 | 16.75±7.80 | < 0.001 | 0.247 | 0.104 |
| Naive/%, $\bar x \pm s$ | 65.03±12.20 | 51.80±13.53 | 55.61±13.01 | 0.041 | 0.177 | 0.574 |
| VAV3+IRF1+B cell/%, $\bar x \pm s$ | 9.46±7.67 | 5.29±1.38 | 12.62±5.16 | 0.124 | 0.329 | < 0.001 |
| Transitional/%, $\bar x \pm s$ | 8.10±2.53 | 4.72±1.64 | 7.04±0.83 | < 0.001 | 0.329 | < 0.001 |
GROUP1, interferon-dominant subtype; GROUP2, B cell activation subtype; GROUP3, endoplasmic reticulum stress subtype; SS, Sjögren syndrome; DN1, double negative 1 B cell; DN2, double negative 2 B cell; Memory, memory B cell; Naive, naïve B cell; Transitional, transitional B cell. |