
恶性肿瘤被认为是一种代谢性疾病,肿瘤细胞的代谢方式比正常细胞更有利于增殖[6]。肿瘤细胞普遍存在瓦博格效应(Warburg effect),即在有氧条件下仍进行高速糖酵解,产生大量乳酸[7]。这种代谢方式虽然效率较低,但却赋予肿瘤细胞很多优势,如增加生物质合成、降低微环境pH值、激活信号通路等。乳酸参与并影响了肿瘤发生发展的多个方面,涉及到肿瘤免疫抑制、肿瘤免疫微环境酸化、肿瘤转移、治疗抵抗和细胞功能等[8]。研究表明,乳酸还可以通过与蛋白质发生共价修饰来调节蛋白质的功能和稳定性,这种修饰被称为乳酸化修饰,是一种新发现的主要发生在组蛋白上的赖氨酸残基表观遗传修饰,可以改变组蛋白的电荷性质,影响染色质的结构和动态,从而调控基因的表达[5, 9]。
代谢重编程与表观遗传重塑作为恶性肿瘤的两大标志,二者密切相关。据报道,在某些情况下,各种代谢改变和表观遗传修饰也会驱动免疫逃逸或阻碍免疫监视,在肿瘤进展中发挥重要作用[10]。蛋白质翻译后修饰(post-translational modification, PTM)作为重要的表观遗传改变,在肿瘤进展中发挥巨大作用[11]。PTM可以影响蛋白质的折叠、稳定性、定位、相互作用、催化活性和信号传导等方面,参与细胞生物学和疾病发生的多个过程。目前已知的PTM有200多种,包括磷酸化、糖基化、泛素化、乙酰化、甲基化等[12]。随着高灵敏度质谱技术的发展,各种代谢物被证明可以通过赖氨酸酰化形式共价修饰蛋白质,包括赖氨酸乙酰化、巴豆酰化、乳酸化、琥珀酰化、丙酰化、丁酰化、丙二酰化、戊二酰化、2-羟基异丁酰化和β-羟基丁酰化[11, 13]。这些修饰可以调节基因表达和细胞内信号通路,并在肿瘤的发生发展中扮演重要角色[14],其中乳酸化修饰作为该家族的新成员,其在不同病理状态下的潜在功能受到了广泛关注,尤其在肿瘤研究领域。本文将聚焦于乳酸化修饰这一新兴领域,系统阐述其调控机制,并重点梳理其在肿瘤代谢、免疫微环境、治疗抵抗等过程中的最新研究进展,以期为开发新的肿瘤诊疗策略提供新的见解。
1 乳酸化进程的发现历程
1.1 乳酸化进程被发现
2019年,一项发表于Nature的成果标志着PTM研究取得重要进展,该研究首次报道了存在于组蛋白上的新型赖氨酸乳酸化修饰[9],通过对人乳腺癌细胞系MCF-7进行组蛋白肽段质谱分析,发现了一种未知的质量偏移,经合成肽段比对、特异性抗体验证及同位素示踪实验,最终证实该修饰源于乳酸,并由糖酵解过程直接供能。研究者进一步发现,在缺氧和M1巨噬细胞极化过程中,组蛋白乳酸化水平呈现动态变化,且其动力学特征不同于经典的乙酰化修饰,这一发现不仅揭示了一种将细胞代谢状态与表观遗传调控直接相连的全新机制,还为乳酸化修饰这一研究方向奠定了基础。
1.2 蛋白质存在广泛的乳酸化进程
继组蛋白乳酸化之后,有研究很快证实乳酸化修饰广泛存在于各类非组蛋白中[15]。在2022年发表于Nature Methods的研究中,研究人员利用合成肽段与理论计算模型,系统解析了乳酸化肽段的质谱裂解规律,首次发现携带乳酸修饰的赖氨酸的多肽在质谱碰撞室中会经历二级断裂,形成链状亚胺离子,然后通过脱氨环化再形成次生碎片,即环状亚胺离子,这一“标志性指纹”极大地提升了鉴定乳酸化修饰位点的准确性与灵敏度。基于此新方法,研究人员对公共蛋白质组学数据进行了重新挖掘,发现了大量新的乳酸化底物,其中许多是糖酵解通路的关键酶(如果糖-1,6-二磷酸醛缩酶),暗示了乳酸化可能存在的自我调节回路[15]。随后的多项研究利用高通量蛋白质组学技术,在健康人肺组织(724个位点)[16]、肝细胞癌组织(2 045个位点)[17-18]、大鼠多器官(1 027个位点)[19]以及胃癌细胞(2 375个位点)[20]中均鉴定出数千个乳酸化位点,且组蛋白修饰仅占很小比例。这些研究均说明乳酸化修饰不仅仅存在于组蛋白上,而是一种普遍存在的PTM现象。与此同时,多个研究也为发现更多更精确的蛋白质乳酸化修饰位点提出新的方式方法[21]。
1.3 蛋白质乳酸化修饰在多个疾病中扮演重要角色
乳酸化修饰一经发现就引起了人们的广泛讨论和研究,随着检测技术的进步,乳酸化修饰在多种生理病理过程中的功能被陆续报道,提示其可能具有多样的生物学功能[22]。例如,在呼吸系统疾病方面,一份亚砷酸盐相关特发性肺纤维化的研究中发现,肌成纤维细胞分泌的乳酸通过促进肺泡上皮细胞中的乳酸化使YTHDF蛋白表达增加,YTHDF1增加了转化生长因子β1(transforming growth factor β1,TGF-β1) 的分泌水平,从而促进了成纤维细胞向肌成纤维细胞的转变[23]。干细胞研究领域,研究者们证明转录因子Glis1诱导干细胞进行多级表观遗传和代谢重塑,从而促进体细胞重编程为多能干细胞。在重编程的早期阶段,Glis1结合并关闭体细胞基因,同时结合糖酵解基因并增强这些基因的转录。随后,增强的糖酵解通量驱动细胞内乙酰辅酶A和乳酸水平上升,进而诱发了多能性基因关键位点的组蛋白乙酰化与乳酸化。这些修饰通过改变染色质的开放状态,有效促进了细胞重编程进程[24]。
2 乳酸化修饰的发生机制
乳酸化修饰是酰化修饰的一种,根据底物蛋白的不同,酰化修饰可大致分为组蛋白酰化修饰和非组蛋白酰化修饰。这种酰化修饰一般发生在赖氨酸残基上,是一种可逆性的过程,需要对应的写入酶让赖氨酸残基发生酰化,消除酶去除酰化修饰,阅读器解释和实现酰化修饰带来的功能改变。以组蛋白乙酰化为例,组蛋白的乙酰化过程由组蛋白乙酰转移酶写入,由组蛋白去乙酰化酶(histone deacetylase, HDAC)家族消除。根据与酵母蛋白的同源性以及催化的结构和生化特征,可以将组蛋白乙酰转移酶划分为三个主要家族:(1)与GCN5蛋白相关的N-乙酰基转移酶;(2)p300/CREB结合蛋白;(3) MYST家族(包括MOZ、Ybf2/Sas3、Sas2、TIP60等蛋白)[30]。
2.1 乳酸来源与活化
2.2 写入酶是乳酸化修饰的催化者
2.3 消除酶是乳酸化修饰的擦除者
2.4 非酶促乳酸化
值得注意的是,有研究报道指出,乙二醛酶代谢的中间产物——乳酰谷胱甘肽,也可以通过非酶催化的反应途径,直接对赖氨酸进行修饰,从而引发乳酸化修饰。D-乳酸主要是通过这种方式生成乳酸化修饰,这种非酶催化的反应机制为我们提供了对乳酸化修饰更加深入的认识[35]。
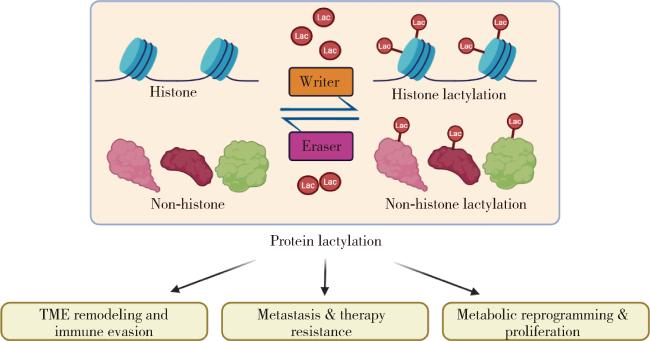
3 乳酸化修饰在恶性肿瘤中的核心功能与机制
{kind=link}
{kind=link}
3.1 驱动免疫抑制与重塑肿瘤微环境
肿瘤微环境中的高水平乳酸是驱动免疫抑制的重要因素,其通过诱导肿瘤细胞自身及浸润免疫细胞发生乳酸化修饰,从多个层面破坏抗肿瘤免疫应答。
在肿瘤细胞中,乳酸化修饰直接上调程序性死亡配体-1等关键免疫检查点分子的表达,介导免疫逃逸。例如,在急性髓系白血病中,STAT5信号驱动的糖酵解亢进导致乳酸累积,通过促进组蛋白乳酸化进而直接上调程序性死亡配体-1的表达[38]。在免疫细胞中,乳酸化修饰则重编程其功能,使其向免疫抑制表型分化。在胶质母细胞瘤中,肿瘤细胞通过PERK-ATF4信号轴重编程单核来源巨噬细胞(monocyte-derived macrophages,MDMs)的代谢,使其糖酵解增强,进而由细胞内乳酸驱动组蛋白乳酸化,直接促进免疫抑制因子白介素-10的转录,从而削弱T细胞功能[39]。此外,在调节性T细胞(regulatory T cells,Treg)和巨噬细胞中,组蛋白H3赖氨酸18位乳酸化(histone H3 lysine 18 lactylation,H3K18la)可提升CD39、CD73等免疫抑制相关基因的启动子活性,进一步破坏免疫平衡[40]。非组蛋白的乳酸化修饰同样参与此过程,例如,乳酸通过修饰膜骨架蛋白MOESIN的赖氨酸72位点,增强其与TGF-β受体Ⅰ及下游SMAD3信号的相互作用,进而促进Treg细胞的生成[41]。
更为复杂的是,乳酸化修饰还能形成自我强化的正反馈环路,持续维持免疫抑制状态。在对胰腺癌的研究中揭示了一个典型环路:乳酸诱导的H3K18la可激活胆固醇代谢酶ACAT2,而ACAT2进一步破坏线粒体功能导致更多乳酸产生;同时,ACAT2通过小细胞外囊泡促使巨噬细胞向更偏向免疫抑制的M2型巨噬细胞极化,共同加剧免疫抑制[42]。
综上所述,乳酸化修饰在肿瘤细胞与免疫细胞间构筑了一个复杂的调控网络,是驱动肿瘤免疫微环境失衡的一个重要环节。
3.2 促进肿瘤转移与治疗抵抗
多向研究表明,乳酸化修饰通过影响基因转录和直接调节非组蛋白功能,参与了肿瘤的侵袭转移过程,并与治疗抵抗的发生相关。
综上所述,现有研究显示乳酸化修饰可能从转录调控和蛋白质功能调节两个层面共同参与肿瘤的转移和耐药过程。
3.3 调控肿瘤细胞代谢与增殖信号
乳酸化修饰与肿瘤代谢重编程之间存在密切的相互促进关系。在胰腺导管腺癌中,有研究报道了一个典型的正反馈回路:糖酵解产生的乳酸导致H3K18la,进而转录激活有丝分裂检查点激酶TTK和BUB1B,而TTK又能反馈性磷酸化并激活糖酵解关键酶——乳酸脱氢酶A(lactate dehydrogenase A,LDHA),从而可能持续驱动瓦博格效应和肿瘤增殖[46]。类似的,纺锤体相关蛋白NUSAP1被报道可与c-Myc蛋白和HIF-1α蛋白结合,增强LDHA启动子活性;而LDHA表达上调和乳酸生成增多,又可引起NUSAP1发生乳酸化修饰并抑制其降解,形成NUSAP1-LDHA-糖酵解-乳酸的反馈环路[47]。这些发现提示,乳酸化修饰可能是维持肿瘤细胞高糖酵解状态的一个潜在环节。
此外,乳酸化修饰还可通过影响关键肿瘤相关蛋白的稳定性与功能,参与增殖信号通路的调节。例如,在结直肠癌研究中,缺氧诱导的糖酵解与β-连环蛋白的乳酸化修饰有关,该修饰可能增强β-连环蛋白的稳定性,与Wnt/β-连环蛋白信号通路的持续激活和细胞恶性行为相关[48]。在肿瘤发生方面,有研究提出,关键抑癌蛋白p53的特定位点乳酸化可能影响其液相分离、DNA结合及转录激活能力,这为理解p53功能失调提供了一个新的视角[27]。在细胞周期调控层面,SIRT3蛋白介导的细胞周期蛋白E2去乳酸化与诱导肝癌细胞死亡相关,反之其乳酸化则可能与细胞周期进程的推动有关[49]。在乳腺癌中,糖酵解来源的乳酸与组蛋白乳酸化支持c-Myc蛋白表达,并可能通过影响下游基因的可变剪接参与肿瘤发生[50]。
蛋白质乳酸化修饰的发现为理解肿瘤代谢与恶性表型之间的内在联系提供了新的分子视角。现有研究表明,乳酸化修饰通过表观遗传调控和非组蛋白功能调节,广泛参与了肿瘤免疫微环境重塑、转移与耐药,以及代谢与增殖信号维持等多个核心生物学过程。这些发现初步揭示了乳酸作为一种代谢物,能够通过共价修饰的方式主动调控细胞功能,将代谢重编程与肿瘤的恶性进展更为直接地联系起来。尽管其特异性功能和作用权重仍需进一步阐明,但针对乳酸化修饰的研究无疑为深入探索肿瘤机制及开发新的干预策略提供了重要的线索和思路。
4 靶向乳酸化修饰的肿瘤治疗策略
鉴于乳酸化修饰在肿瘤中的潜在作用,探索针对该修饰的治疗策略已成为一个新兴方向。目前的策略主要围绕三个层面展开:调控乳酸代谢以削减修饰前体、直接干预修饰酶系统,以及开发特异性阻断工具。
4.1 源头干预:调控乳酸代谢
通过抑制乳酸生成或阻断其外排来降低肿瘤微环境中的乳酸水平是间接影响乳酸化修饰的基础策略,这种策略可以简单分为抑制乳酸生成和阻断乳酸转运体单羧酸转运蛋白(monocarboxylate transporters,MCT)[8]。
LDHA作为催化丙酮酸生成乳酸的关键酶,在多种肿瘤中高表达。临床前研究证实,抑制LDHA不仅可有效遏制肿瘤生长与转移,还能增强传统化疗的敏感性[2, 29, 51-54]。目前已开发出多种类型的LDHA小分子抑制剂,如司替戊醇、草酸盐、棉酚等,其中司替戊醇在细胞和动物模型中显示出与化疗的协同增效作用[28, 55]。然而,该策略面临挑战:LDHA在正常代谢中的作用很关键,但其系统性抑制可能引发NAD+耗竭所导致的代谢紊乱,或引起细胞外基质重塑促进转移等副作用[56]。尽管存在上述挑战,但靶向LDHA在增强传统肿瘤疗法的敏感性方面仍具有明确的潜力,因此,优化用药策略并推进临床转化以验证其疗效成为下一步研究的关键方向。
4.2 直接靶向:干预修饰酶系统
直接调控乳酸化修饰的写入酶和消除酶,是比降低乳酸水平更为精准的干预策略。然而,目前已鉴定的相关酶类(如p300/CREB结合蛋白和HDAC家族)通常缺乏对乳酸化修饰的特异性,这既是对当前干预策略的主要挑战,也揭示了其作用的复杂性。
理论上,激活去乳酸化酶(如HDAC1-3)是降低乳酸化水平的合理策略,但这一思路与临床观察相悖:广泛使用的HDAC抑制剂,如伏立诺他(vori-nostat)等,反而表现出抗癌活性[59-62],几种脱乙酰酶抑制剂已被美国食品和药物管理局批准用于治疗淋巴瘤和骨髓瘤。这一矛盾现象揭示了HDAC家族功能的复杂性:它们并非乳酸化的特异性消除酶,同时对乙酰化等多种酰化修饰具有活性。因此,HDAC抑制剂的净效应是重塑整体的蛋白质酰化修饰谱,其抗癌作用可能源于这种更广泛的表观遗传重编程,而非单纯提升乳酸化水平。这一发现具有双重意义:一方面,解释了基于HDAC抑制剂的疗法已被美国食品和药物管理局批准用于治疗淋巴瘤和骨髓瘤,可被视为一种通过广泛调控酰化修饰谱来间接影响乳酸化水平的“粗调”策略[59-62];另一方面,提示未来的发展方向是开发能特异性靶向HDAC家族去乳酸化功能的新型调控分子,以减少其对整体酰化修饰谱的脱靶效应。
4.3 精准探索:新兴的特异性干预策略
5 总结
乳酸化修饰的发现为理解细胞代谢状态与基因表达调控之间的精确联系提供了关键分子桥梁。本文系统梳理了乳酸化修饰从发现、机制到在肿瘤中的多功能作用及靶向策略,着重阐述了其如何将表观遗传调控与代谢重编程相联系,并参与肿瘤发生、免疫逃逸及治疗抵抗等过程的潜在机制。然而,作为新兴领域,乳酸化研究在技术、机制认知及临床转化方面仍面临诸多挑战,同时也蕴含着广阔机遇。(1)在技术层面,乳酸化修饰的精准鉴定仍是该领域发展的基石,也是当前的主要瓶颈。目前,乳酸化修饰位点的鉴定主要依赖于基于质谱的蛋白质组学技术,然而,该方法对样本量要求高、耗时且成本高昂,对低丰度或低覆盖度的修饰位点检测能力有限。新技术的突破有望推动更多功能性乳酸化蛋白的发现,从而深化我们对乳酸化修饰的理解。(2)在分子机制上,乳酸化写入酶与消除酶的特异性及调控网络是理解其功能复杂性的核心。目前已知的关键酶如p300/CREB结合蛋白和HDAC家族,均非乳酸化修饰的特异性酶,它们同时广泛参与乙酰化等多种酰化修饰的调控。这不仅解释了为何临床上使用广谱HDAC抑制剂能发挥抗癌作用(其净效应是改变整体酰化修饰谱,而非单纯升高乳酸化水平),也提示细胞内可能存在一个由相同写入酶/消除酶调控的、乳酸化与乙酰化等修饰相互对话的精密网络。一种可能的调控模式是,作为糖酵解分支产物的乳酸和乙酰辅酶A,可能通过竞争相同的修饰酶,根据细胞代谢状态(如缺氧/富氧)动态调控组蛋白的乳酸化与乙酰化水平,从而将不同的代谢信号转化为特定的表观遗传指令。(3)研究证据表明,乳酸化调控具有显著的位点特异性。例如,在肝癌中,去甲泽拉木醛处理虽降低总乳酸水平,但仅特异性降低了组蛋白H3赖氨酸9位和56位的乳酸化,而对14位和23位的赖氨酸乳酸化则无影响[63]。这强烈提示,不同位点的乳酸化可能由不同的、具有底物特异性的写入酶催化。非组蛋白如Vps34被TIP60乳酸化进而调控自噬的案例[64]进一步支持了“不同底物甚至同一底物的不同位点可能由特定酶调控”的假说。因此,未来研究的重点之一将是系统性地鉴定和验证这些具有高度特异性的乳酸化修饰酶。(4)在研究方法上,当前研究多通过调控乳酸代谢(如使用糖酵解或MCT抑制剂)来间接影响乳酸化水平,这使得准确区分乳酸本身的信号功能与其作为前体形成的乳酸化修饰的功能变得困难。因此,开发能够直接、特异地干预某种蛋白的特定乳酸化修饰,而不全局改变乳酸水平或其他修饰的工具(如MRE11乳酸化阻断肽[30]),对于精确解析乳酸化的独立功能至关重要,这类工具也将为开发副作用更小的靶向治疗提供新途径。(5)从更广阔的视角看,乳酸化修饰的发现,为代谢重编程与表观遗传重塑这两大肿瘤标志提供了全新的直接连接点,这提示乳酸不仅是代谢废物,其本身也可能作为一种活跃的调控分子,通过乳酸化修饰直接参与染色质和功能蛋白的调节,从而影响细胞命运。未来的研究不仅需要绘制完整的乳酸化修饰图谱,更需要深入揭示其在不同肿瘤背景下的特异性功能,并积极推动特异性靶向策略(如MRE11乳酸化阻断肽)向临床转化。乳酸化研究是一个新兴的研究领域,随着检测技术的革新、机制研究的深入和靶向策略的精准化,对乳酸化修饰的探索将继续深化我们对肿瘤代谢-表观遗传交叉对话的理解,并可能为肿瘤诊疗新策略的开发提供新的参考。