
1 材料与方法
1.1 主要材料及试剂
1.2 原代rBMSCs的提取与培养
1.3 rBMSCs成神经诱导
1.4 rBMSCs成骨诱导
1.5 miRNA转染
1.6 RNA提取、逆转录与qRT-PCR
表1 qRT-PCR引物序列Table 1 Primer sequences used for qRT-PCR |
| Gene | Forward primer (5′ to 3′) | Reverse primer (5′ to 3′) |
| U6 | CGCTTCGGCAGCACATATAC | CGAATTTGCGTGTCATCCTT |
| miR-488-5p | GGCACCCAGATAATGGCAC | AGTGCAGGGTCCGAGGTATT |
| Gapdh | AGGTCGGTGTGAACGGATTT | GAACTTGCCGTGGGTAGAGT |
| *Map2 | ACAGCAACAAGTGGTGAATCAG | GGAGGATGGAGGAAGGTCTTG |
| *Ng2 | TTCTCACACAGAGGAGCCC | CACTCAAGCTCTGGCTGCT |
| *Tubb3 | CAGATGCTGGCCATTCAGAGTAAG | TGTTGCCGATGAAGGTGGAC |
| *Ngf | TGCCAAGGACGCAGCTTTC | TGAAGTTTAGTCCAGTGGGCTTCAG |
| *Nes | TGAACAAGAGACCCAACAAACAC | TTCCAAGAGGCTTCGGTAACT |
| *S100 | CTGTCAAGAACCTGCTCCGA | AGTGGGCATGGAACACATTGA |
| #Runx2 | GCCTTCAAGGTTGTAGCCCT | TGAACCTGGCCACTTGGTTT |
| #Alp | CATGGTGAGTGACACGGACA | CCATGACGTGGGGGATGTAG |
| #Ocn | CCGTTTAGGGCATGTGTTGC | TTTCGAGGCAGAGAGAGGGA |
| #Bmp2 | CGGGAACAAATGCAGGAAGC | AAGGACATTCCCCATGGCAG |
| #Col1a1 | CCCCAGCCGCAAAGAGTCTA | CAGCTGACTTCAGGGATGTCTTC |
| #Opn | CCAGCCAAGGACCAACTACA | AGTGTTTGCTGTAATGCGCC |
* neurogenic-related genes; # osteogenic-related genes; qRT-PCR, quantitative real-time polymerase chain reaction; Map2, microtubule-associated protein 2; Ng2, neural/glial antigen 2; Tubb3, Class Ⅲ β-tubulin; Ngf,nerve growth factor;Nes, nestin; S100, soluble protein-100; Runx2, runt-related transcription factor 2; Alp, alkaline phosphatase; Ocn, osteocalcin; Bmp2, bone morphogenetic protein 2; Col1a1, collagen type Ⅰ alpha 1 chain; Opn, osteopontin. |
1.7 蛋白免疫印迹实验
1.8 细胞免疫荧光实验
1.9 ALP染色与定量实验
1.10 茜素红矿化结节染色(alizarin red staining,ARS)与半定量实验
1.11 大鼠颅骨缺损模型构建与实验分组
1.12 显微计算机断层扫描术(micro computed tomography,micro-CT)与骨参数分析
1.13 组织形态学分析
1.14 统计学分析
2 结果
2.1 miR-488-5p在rBMSCs成神经分化过程中表达上调
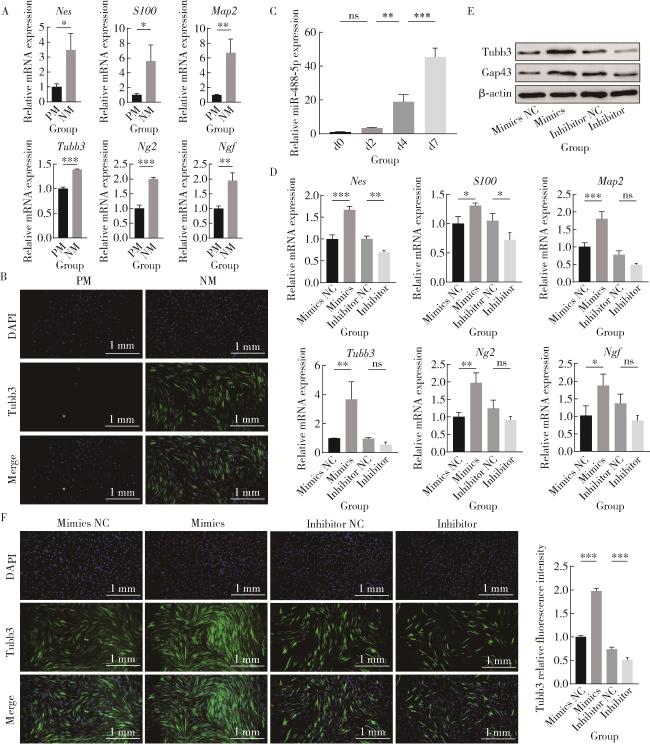
图1 miR-488-5p促进rBMSCs神经向分化Figure 1 miR-488-5p promotes neurogenic differentiation of rBMSCs A, mRNA expression levels of neurogenic-related genes Nes, S100, Map2, Ng2, Tubb3, Ngf on day 12 of neurogenic differentiation in the NM and PM groups as detected by qRT-PCR; B, immunofluorescence detection of Tubb3 in the NM and PM groups; C, miR-488-5p expression levels on days 0, 2, 4 and 7 of neurogenic differentiation as detected by qRT-PCR; D, mRNA expression levels of neurogenic-related genes Nes, S100, Map2, Ng2, Tubb3, Ngf on day 12 of neurogenic differentiation as detected by qRT-PCR; E, protein expression levels of Tubb3 and Gap43 as detected by Western blot; F, immunofluorescence detection of Tubb3 and quantification of Tubb3 fluorescence intensity. ns, not significant. * P < 0.05, * * P < 0.01, * * * P < 0.001. PM, proliferation medium; NM, neurogenic medium; Mimics NC, negative control for miR-488-5p mimics; Inhibitor NC, negative control for miR-488-5p inhibitor; Nes, nestin; S100, soluble protein-100; Map2, microtubule-associated protein 2; Ng2, neural/glial antigen 2; Tubb3, class Ⅲ β-tubulin; Ngf, nerve growth factor; Gap43, growth associated protein 43;DAPI, 4', 6-diamidino-2-phenylindole; rBMSCs, rat bone marrow mesenchymal stem cells; qRT-PCR, quantitative real-time polymerase chain reaction. |
2.2 miR-488-5p促进rBMSCs成神经分化
2.3 miR-488-5p在rBMSCs成骨分化过程中表达上调
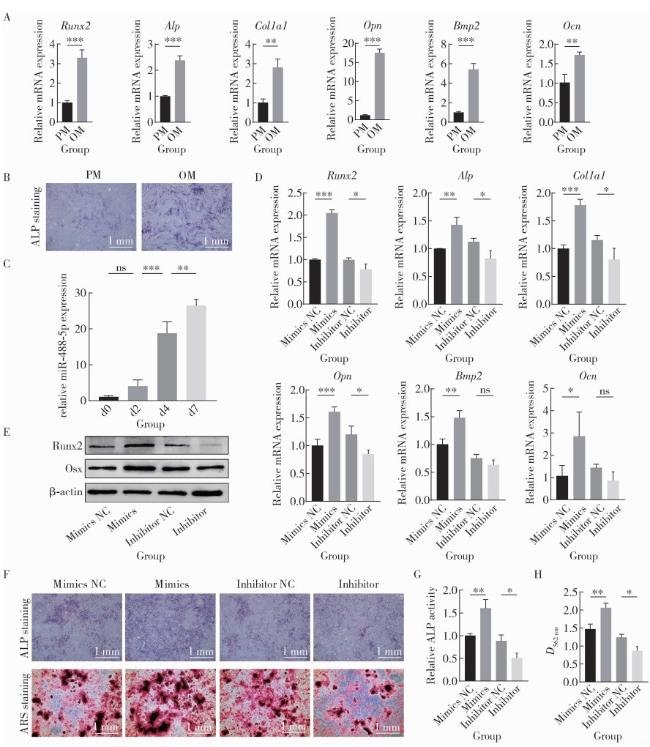
图2 miR-488-5p促进rBMSCs成骨向分化Figure 2 miR-488-5p promotes osteogenic differentiation of rBMSCs A, mRNA expression levels of osteogenic-related genes Runx2, Alp, Col1a1, Opn, Bmp2, Ocn on day 7 of osteogenic differentiation in the OM and PM groups as detected by qRT-PCR; B, ALP staining on day 7 of osteogenic differentiation in the OM and PM groups; C, miR-488-5p expression levels on days 0, 2, 4 and 7 of osteogenic differentiation as detected by qRT-PCR; D, mRNA expression levels of osteogenic-related genes Runx2, Alp, Col1a1, Opn, Bmp2, Ocn on day 7 of osteogenic differentiation as detected by qRT-PCR; E, protein expression levels of Runx2 and Osx as detected by Western blot; F, ALP staining on day 7 and ARS staining on day 21 of osteogenic differentiation; G, ALP activity on day 7 of osteogenic differentiation; H, quantification of mineralization by measuring ARS absorbance at 562 nm. ns, not significant. * P < 0.05, * * P < 0.01, * * * P < 0.001. PM, proliferation medium; OM, osteogenic medium; d, day; Mimics NC, negative control for miR-488-5p mimics; Inhibitor NC, negative control for miR-488-5p inhibitor; Runx2, Runt-related transcription factor 2; ALP, alkaline phosphatase; Ocn, Osteocalcin; Bmp2, bone morphogenetic protein 2; Col1a1, collagen type Ⅰ alpha 1 chain; Opn, osteopontin; ARS, alizarin red S; rBMSCs, rat bone marrow mesenchymal stem cells; qRT-PCR, quantitative real-time polymerase chain reaction. |
2.4 miR-488-5p促进rBMSCs成骨分化
2.5 过表达miR-488-5p促进大鼠临界尺寸颅骨缺损神经化组织工程骨形成
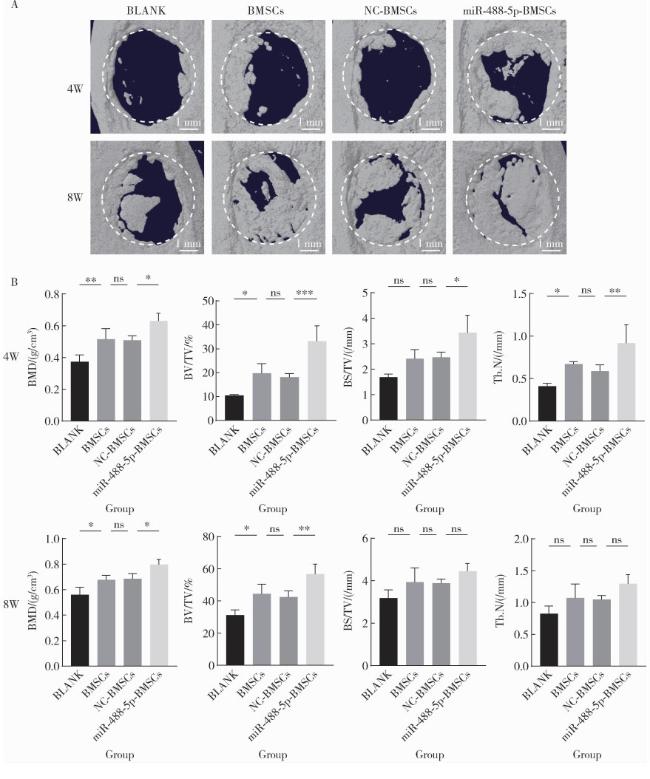
图3 大鼠颅骨缺损术后4周和8周micro-CT影像学分析Figure 3 micro-CT imaging analysis of rat calvarial defects at 4 and 8 weeks after operation A, 3D reconstructed images after micro-CT scanning showing bone defect areas at both 4 and 8 weeks after operation; B, quantitative comparisons of BMD, BV/TV, BS/TV, Tb.N at both 4 and 8 weeks after operation. ns, not significant. * P < 0.05, * * P < 0.01, * * * P < 0.001. W, weeks after operation. BLANK, the control group treated with GelMA only; BMSCs, the group treated with GelMA and normal rBMSCs; NC-BMSCs, the group treated with GelMA and rBMSCs transfected with negative control (NC) for miR-488-5p mimics; miR-488-5p-BMSCs, the group treated with GelMA and rBMSCs transfected with miR-488-5p mimics; BMD, bone mineral density; BV/TV, bone volume/total volume; BS/TV, bone surface area/total volume; Tb.N, trabecular number; 3D, 3-dimensional; BMSCs, bone marrow mesenchymal stem cells. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
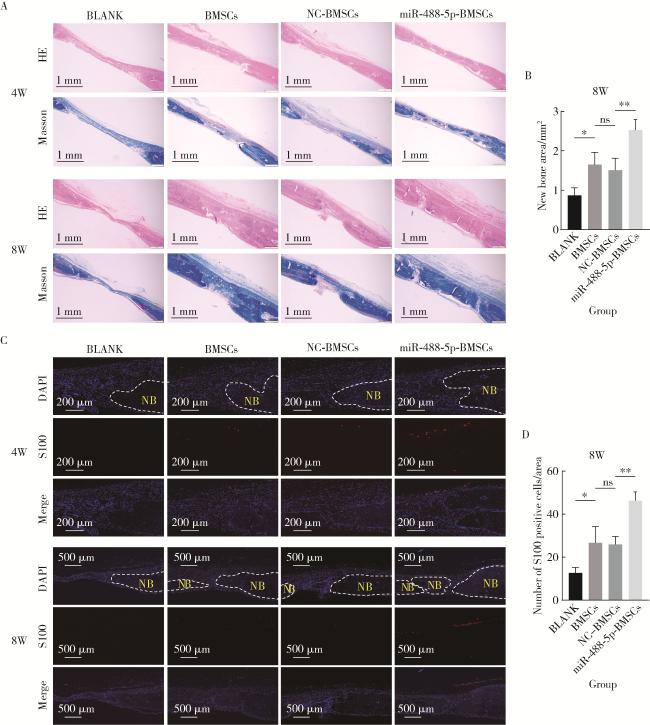
图4 大鼠颅骨缺损术后4周和8周组织形态学分析Figure 4 Histomorphological analysis of rat calvarial defects at 4 and 8 weeks after operation A, HE staining and Masson staining; B, quantitative measurement of new bone area in Masson staining at 8 weeks after operation; C, immunofluorescence detection of S100 protein; D, analysis of the number of S100-positive cells in immunofluorescence staining at 8 weeks after operation. HE, Hematoxylin-Eosin; NB, new bone; S100, soluble protein-100; DAPI, 4', 6-diamidino-2-phenvlindole; W, weeks after operation; BLANK, the control group treated with GelMA only; BMSCs, the group treated with GelMA and normal rBMSCs; NC-BMSCs, the group treated with GelMA and rBMSCs transfected with negative control (NC) for miR-488-5p mimics; miR-488-5p-BMSCs, the group treated with GelMA and rBMSCs transfected with miR-488-5p mimics; BMSCs, bone marrow mesenchymal stem cells. ns, not significant; * P < 0.05, * * P < 0.01. |