北京大学学报(医学版) ›› 2020, Vol. 52 ›› Issue (2): 214-220. doi: 10.19723/j.issn.1671-167X.2020.02.004
角蛋白18与剪切因子PSF相互作用介导PSF的膜易位并维持髓系白血病细胞的化疗敏感性
任思楣1,2,△( ),龙璐瑶1,2,许成山1
),龙璐瑶1,2,许成山1
- 1. 北京医院 国家老年医学中心 国家卫生健康委员会临床检验中心 中国医学科学院老年医学研究院,北京 100730
2. 中国医学科学院北京协和医学院研究生院,北京 100730
Interaction between PSF and cytokeratin 18 mediates PSF relocation to cell membrane and maintains chemosensitivity of myeloid leukemia
Si-mei REN1,2,△(),Lu-yao LONG1,2,Cheng-shan XU1
- 1. National Center for Clinical Laboratories, Beijing Hospital, National Center of Gerontology; Institute of Geriatric Medicine, Chinese Academy of Medical Sciences, Beijing 100730, China
2. Graduate School of Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing 100730, China
摘要:
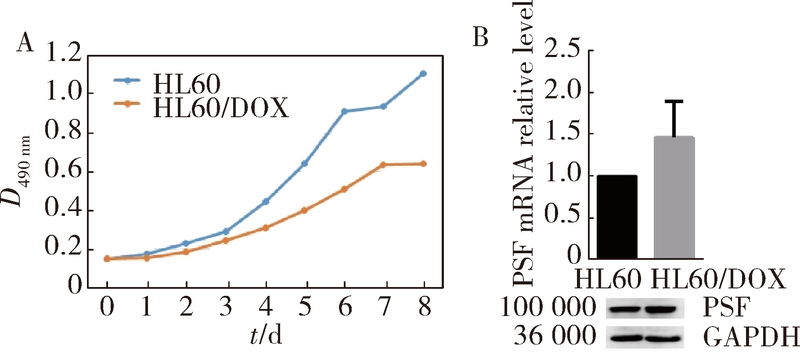
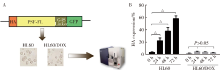
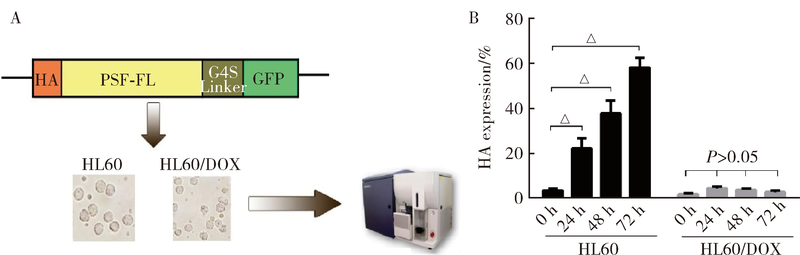
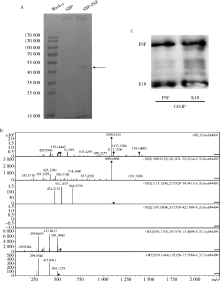
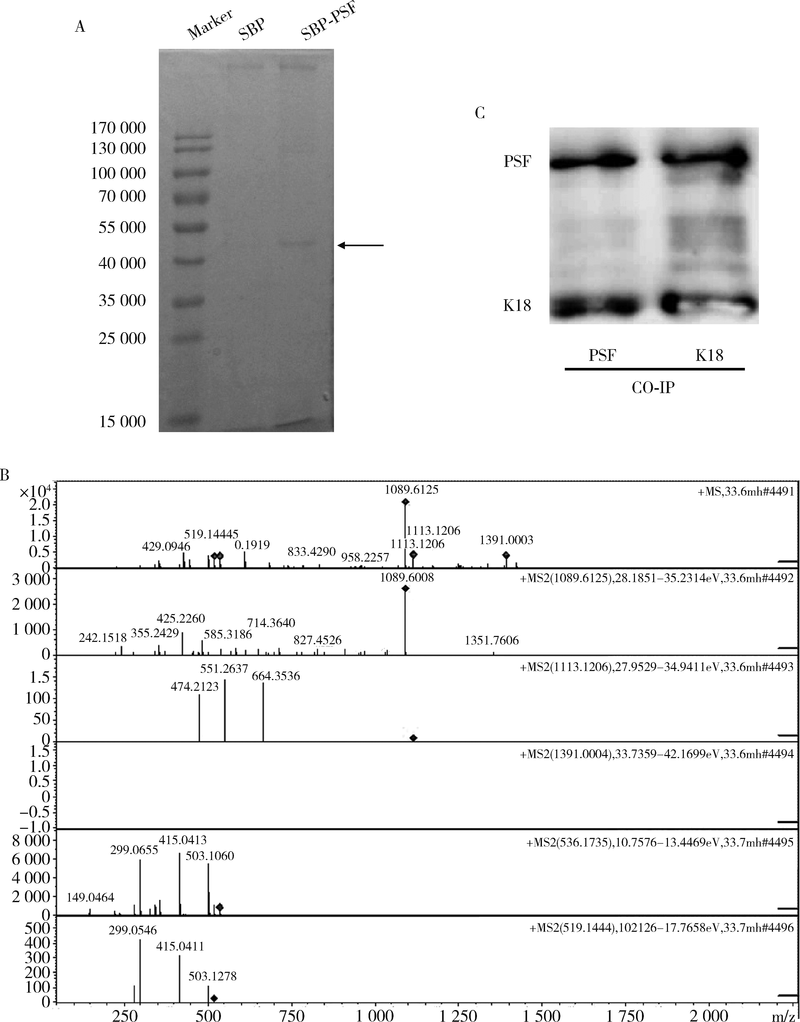
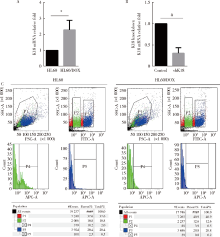
目的 鉴定多嘧啶结合蛋白相关的剪切因子(polypyrimidine tract-binding protein-associated splicing factor, PSF)在髓系白血病细胞内的伴侣蛋白,探讨PSF在敏感HL60细胞和耐药的HL60/DOX细胞中易位细胞膜的模式和机制.方法: 通过脂质体瞬时转染PSF真核表达载体过表达PSF,进一步结合流式细胞术在转染后24 h,48 h,72 h检测PSF在细胞膜上的表达水平.构建链亲和素结合肽(streptavidin binding peptide,SBP)和PSF的融合表达载体,转染载体,过表达SBP-PSF融合蛋白.通过链亲和素磁珠共沉淀法和质谱分析,鉴定PSF在细胞内的伴侣蛋白.在慢病毒载体中插入角蛋白18 (cytokeratin 18,K18)干扰序列,转染293T细胞制备病毒液.用慢病毒感染HL60和HL60/DOX细胞,获得稳定干扰K18的细胞株.结合流式细胞术检测干扰K18的HL60和HL60/DOX细胞中细胞膜上PSF的表达水平,由此证实CK18在HL60和HL60/DOX细胞中协同转运PSF易位细胞膜机制的异同.结果: 瞬时过表达PSF后的24 h,48 h,72 h连续3 d检测细胞膜PSF表达水平,HL60敏感株细胞膜上PSF表达率分别为22.4%±3.5%, 37.9%±6.0%, 58.3%±8.8%;耐药株HL60/DOX细胞膜上PSF的表达率分别为4.7%±0.5%, 3.9%±0.6%, 2.9%±0.6%;各时间点下敏感株和耐药株的差异有统计学意义,P<0.01.免疫共沉淀和质谱证实K18为PSF在细胞内的伴侣蛋白.干扰K18的表达后,再次分析细胞膜PSF表达,发现PSF在敏感株细胞膜上的表达水平由原来的48.9%±5.4% 降至6.2%±1.0%;在耐药株细胞膜上的表达水平由9.11%±1.2%降至2.21%±0.51%.结论: K18是PSF在细胞内的伴侣蛋白,在敏感细胞中,K18与PSF相互作用可帮助PSF向细胞膜转运;而在耐药株中,由于该功能障碍导致PSF在胞内发生积累从而介导耐药性.
中图分类号:
- R557.1


| [1] | Kulkoyluoglu-Cotul E, Smith BP, Wrobel K , et al. Combined targeting of estrogen receptor Alpha and XPO1 prevent akt activation, remodel metabolic pathways and induce autophagy to overcome tamoxifen resistance[J]. Cancers (Basel), 2019,11(4):479. |
| [2] | Ren S, She M, Li M , et al. The RNA/DNA-binding protein PSF relocates to cell membrane and contributes cells' sensitivity to antitumor drug, doxorubicin[J]. Cytometry A, 2014,85(3):231-241. |
| [3] | Knott GJ, Bond CS, Fox AH . The DBHS proteins SFPQ, NONO and PSPC1: A multipurpose molecular scaffold[J]. Nucleic Acids Res, 2016,44(9):3989-4004. |
| [4] | Gao Z, Chen M, Tian X , et al. A novel human lncRNA SANT1 cis-regulates the expression of SLC47A2 by altering SFPQ/E2F1/HDAC1 binding to the promoter region in renal cell carcinoma[J]. RNA Biol, 2019,16(7):940-949. |
| [5] | de Silva HC, Lin MZ, Phillips L , et al. IGFBP-3 interacts with NONO and SFPQ in PARP-dependent DNA damage repair in triple-negative breast cancer[J]. Cell Mol Life Sci, 2019,76(10):2015-2030. |
| [6] | Takayama K I, Suzuki T, Fujimura T , et al. Dysregulation of spliceosome gene expression in advanced prostate cancer by RNA-binding protein PSF[J]. Proc Natl Acad Sci USA, 2017,114(39):10461-10466. |
| [7] | Yarosh CA, Iacona JR, Lutz CS , et al. PSF: nuclear busy-body or nuclear facilitator?[J]. Wiley Interdiscip Rev RNA, 2015,6(4):351-367. |
| [8] | Izumi H, Mccloskey A, Shinmyozu K , et al. p54nrb/NonO and PSF promote U snRNA nuclear export by accelerating its export complex assembly[J]. Nucleic Acids Res, 2014,42(6):3998-4007. |
| [9] | Duan Y, Sun Y, Zhang F , et al. Keratin K18 increases cystic fibrosis transmembrane conductance regulator (CFTR) surface expression by binding to its C-terminal hydrophobic patch[J]. J Biol Chem, 2012,287(48):40547-40559. |
| [10] | Chen J, Ouyang H, An X , et al. Vault RNAs partially induces drug resistance of human tumor cells MCF-7 by binding to the RNA/DNA-binding protein PSF and inducing oncogene GAGE6[J]. PLoS One, 2018,13(1):e0191325. |
| [11] | Wang G, Cui Y, Zhang G , et al. Regulation of proto-oncogene transcription, cell proliferation, and tumorigenesis in mice by PSF protein and a VL30 noncoding RNA[J]. Proc Natl Acad Sci USA, 2009,106(39):16794-16798. |
| [12] | Wang XT, Xia QY, Ni H , et al. SFPQ/PSF-TFE3 renal cell carcinoma: a clinicopathologic study emphasizing extended morphology and reviewing the differences between SFPQ-TFE3 RCC and the corresponding mesenchymal neoplasm despite an identical gene fusion[J]. Hum Pathol, 2017,63:190-200. |
| [13] | Ishikawa N, Nagase M, Takami S , et al. Xp11.2 translocation renal cell carcinoma with SFPQ/PSF-TFE3 fusion gene: A case report with unusual histopathologic findings[J]. Pathol Res Pract, 2019,215(9):152479. |
| [14] | Ha K, Takeda Y, Dynan WS . Sequences in PSF/SFPQ mediate radioresistance and recruitment of PSF/SFPQ-containing complexes to DNA damage sites in human cells[J]. DNA Repair (Amst), 2011,10(3):252-259. |
| [15] | Zou Y, He L, Wu CH , et al. PSF is an IbeA-binding protein contributing to meningitic Escherichia coli K1 invasion of human brain microvascular endothelial cells[J]. Med Microbiol Immunol, 2007,196(3):135-143. |
| [16] | Toivola DM, Boor P, Alam C , et al. Keratins in health and disease[J]. Curr Opin Cell Biol, 2015,32:73-81. |
| [17] | Ramot Y, Zlotogorski A . Keratins: the hair shaft's backbone revealed[J]. Exp Dermatol, 2015,24(6):416-417. |
| [18] | Lee S Y, Kim S, Lim Y , et al. Keratins regulate Hsp70-mediated nuclear localization of p38 mitogen-activated protein kinase[J/OL]. J Cell Sci, 2019, 9(2019-09-26)[2019-11-09]. |
| [19] | Zhang B, Wang J, Liu W , et al. Cytokeratin 18 knockdown decreases cell migration and increases chemosensitivity in non-small cell lung cancer[J]. J Cancer Res Clin Oncol, 2016,142(12):2479-2487. |
| [20] | Shi R, Wang C, Fu N , et al. Downregulation of cytokeratin 18 enhances BCRP-mediated multidrug resistance through induction of epithelial-mesenchymal transition and predicts poor prognosis in breast cancer[J]. Oncol Rep, 2019,41(5):3015-3026. |
| No related articles found! |
|
||
