
1 材料与方法
1.1 主要材料与试剂
1.2 DPSCs及3T3细胞的培养
1.3 细胞膜囊泡的制备及检测
1.4 siRNA的加载
1.5 流式细胞术测定
1.6 细胞毒性测定
1.7 RNA提取和实时荧光定量聚合酶链反应(quantitative real-time polymerase chain reaction, qRT-PCR)测定
表1 实时荧光定量PCR引物序列Table 1 Quantitative real-time PCR primer sequence |
| Gene | Forward primer (5′ to 3′) | Reverse primer (5′ to 3′) |
| TNF-α | AGCAAGGACAGCAGAGGA | GGGGAGAGAGGGTGGAG |
| IL-1β | TGTGCTGAATGTGGACTCA | ACAAAAGGGCTGGGGAT |
| IL-6 | TGTGTGAAAGCAGCAAAGA | ACCAGGCAAGTCTCCTCA |
| GAPDH | GCCAACGTGTCAGTGGTG | AAGGTGGAGGAGTGGGTGT |
PCR, polymerase chain reaction; TNF-α, tumor necrosis factor-alpha; IL-1β, interleukin-1 beta; IL-6, interleukin-6; GADPH, glyceraldehyde-3-phosphate dehydrogenase. |
1.8 酶联免疫吸附试验(enzyme-linkel immunosorbent assay, ELISA)测定
1.9 统计学分析
2 结果
2.1 细胞膜囊泡的制备
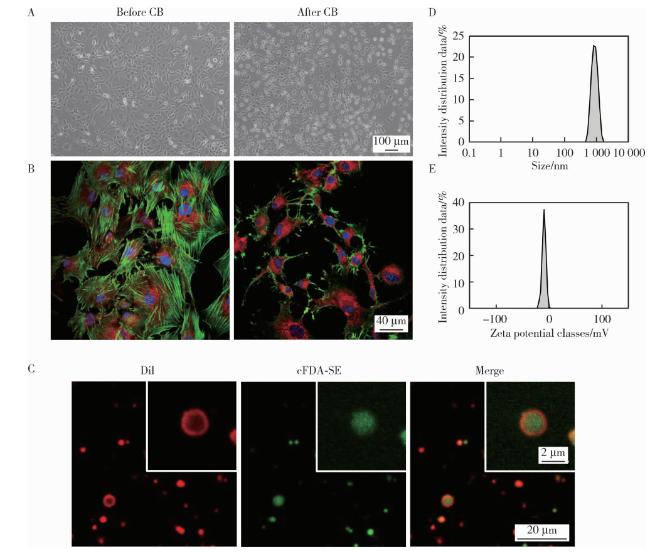
图1 CMVs的制备及表征Figure 1 Preparation and characterization of CMVs A, morphological changes of 3T3 cells without treatment and after 30 min of cytochalasin B exposure under a light microscope (scale bar=100 μm); B, alterations of cell membrane and cytoskeleton in untreated and CB-treated (30 min) 3T3 cells observed by confocal microscopy (red: DiI, green: phalloidin, blue: DAPI; scale bar = 40 μm); C, confocal microscopy images of CMVs (red: DiI, green: cFDA-SE, scale bar = 20 μm) and magnified view (red: DiI, green: cFDA-SE, scale bar = 2 μm); D, size distribution of CMVs; E, zeta potential distribution of CMVs. DAPI, 4', 6-diamidino-2-phenylindole; cFDA-SE, 5-(and-6)-carboxyfluorescein diacetate succinimidyl ester; DiI, 1, 1'-dioctadecyl-3, 3, 3', 3'-tetramethylindocarbocyanine perchlorate; CMVs, cell membrane vesicles; CB, cytochalasin B. |
2.2 siRNA加载
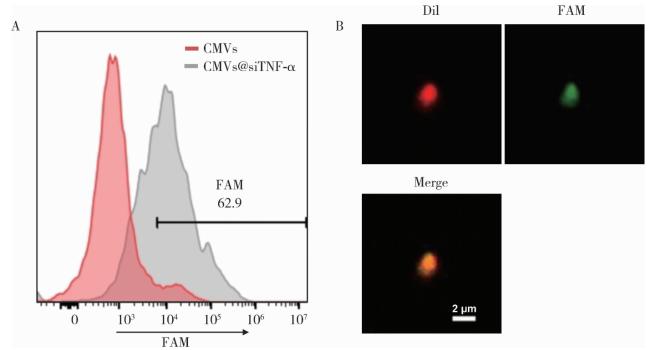
图2 CMVs对siTNF-α的加载Figure 2 Loading of siTNF-α into CMVs A, flow cytometry analysis of CMVs with or without loading of FAM-siTNF-α; B, confocal microscopy images of CMVs@FAM-siTNF-α (red, DiI; green, FAM; scale bar=2 μm). DiI, 1, 1′-dioctadecyl-3, 3, 3', 3'-tetramethylindocarbocyanine perchlorate; CMVs, cell membrane vesicles; FAM, fluorescein amidite; siTNF-α, small interfering RNA targeting tumor necrosis factor-alpha. |
2.3 CMVs@siTNF-α在细胞内的表达和作用
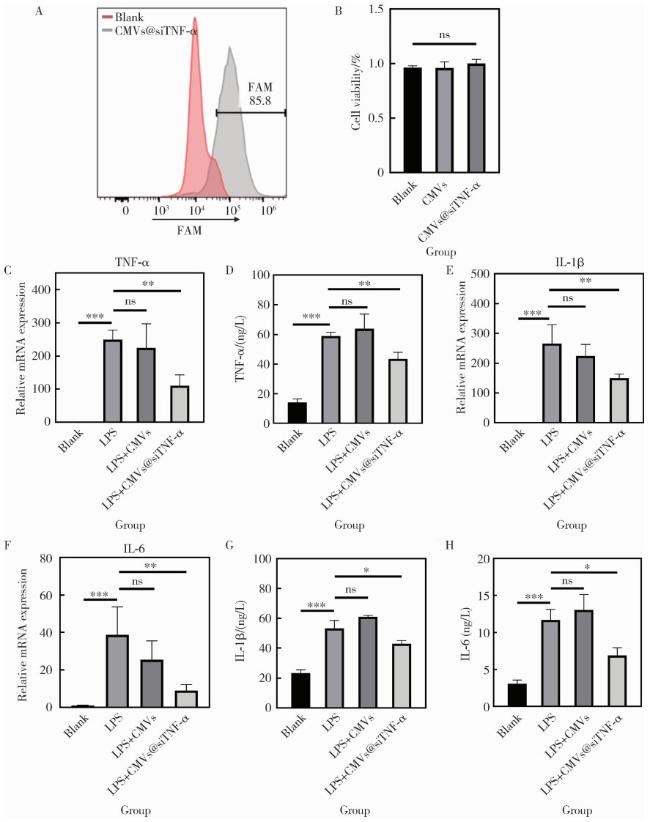
图3 CMVs@siTNF-α在DPSCs中的转运与抗炎效果Figure 3 Intracellular delivery and anti-inflammatory effects of CMVs@siTNF-α A, flow cytometry analysis of DPSCs with or without transfection of CMVs@siTNF-α; B, relative cell viability of DPSCs after treatment with CMVs or CMVs@siTNF-α, measured by CCK-8 assay; C, E, F, mRNA expression levels of TNF-α, IL-1β, and IL-6 in DPSCs under different treatments (blank, LPS, LPS+CMVs, or LPS+CMVs@siTNF-α), determined by qRT-PCR; D, G, H, protein secretion levels of TNF-α, IL-1β, and IL-6, assessed by ELISA. *P < 0.05; * *P < 0.01; * * *P0.001; ns, no significance. CMVs, cell membrane vesicles; siTNF-α, small interfering RNA targeting tumor necrosis factor-alpha; FAM, fluorescein amidite; LPS, lipopolysaccharide; TNF-α, tumor necrosis factor-alpha; IL-1β, interleukin-1 beta; IL-6, interleukin-6; DPSCs, dental pulp stem cells; qRT-PCR, quantitative real-time polymerase chain reaction; ELISA, enzyme-linked immunosorbent assay. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}